
Abstract
This study describes the introduction and spread of avian influenza A (H5N1) subtype in Croatia. Seventeen isolates were identified during the period from October 2005 to March 2006, all originating from wild birds. The full-length nucleotide sequence analysis of the hemagglutinin (HA) gene of seven representative isolates revealed that three distinct genetic strains involved in the outbreaks, implicating at least three independent introductions of the virus into Croatia during a relatively short period of time. All three genetic strains belonged to clade 2.2 (Qinghai-like viruses) and each strain displayed significant similarity to concurrent H5N1 viruses from other European countries. The dominant strain of the virus was present in all four affected areas and in all three bird species (mute swan, mallard, and black-headed gull), indicating cross-species transmission of the virus. Two other genetic strains were found, together with the dominant strain, only in a marsh at the Adriatic coast during late February and early March 2006, which could be associated with frozen water surfaces in the continental part of Croatia as well as in Eastern Europe in early 2006 and the movement of birds toward warmer areas. This is also the first isolation of highly pathogenic avian influenza virus of H5N1 subtype from apparently healthy black-headed gulls.
Introduction
Asian HPAIV of H5N1 subtype apparently emerged in domestic poultry in China already in 1996 (Xu et al. 1999) and circulated locally with sporadic spread to neighboring Hong Kong (Sims et al. 2005). The situation changed dramatically between 12 December, 2003 and 27 January, 2004 when eight countries in South-East Asia first reported the presence of the virus in poultry (Capua and Alexander 2004). The widespread presence of HPAI H5N1 viruses in poultry in the Far East inevitably resulted in spill-over into wild bird populations in May 2005 (Alexander and Capua 2008). Probably both wild bird migrations and the movement of poultry were responsible for further spread of the virus westward (Alexander and Capua 2008), and there is strong evidence that wild birds could spread the virus long distances (Kilpatrick et al. 2006, Normile 2006).
Early isolates from wild mute swans (Cygnus olor) were obtained in Croatia already in October 2005 (OIE 2009). These infected swans were a forerunner of the apparent importance of these birds in the spread of the HPAIV H5N1, and from January to April 2006, wild mute swans and other wild birds were shown to be infected in numerous countries in Europe, Caucasus, and Middle East (Alexander and Capua 2008). Genome analysis of HPAI H5N1 viruses from wild and domestic birds and humans in Europe and Africa show their relationship with the isolates from wild birds at the Qinghai Lake in China. This Euro-African lineage, which was the cause of several fatal human infections in Middle East and Africa, has been introduced at least three times into the European-African region and has split into three distinct, independently evolving sublineages (Salzberg et al. 2007).
Apart from the initial introduction and finding of the HPAI H5N1 virus in wild swans in October 2005, the virus has been found in several wild bird species in four major areas in Croatia until late March 2006. In this study we described details of the HPAI H5N1 virus infection in wild birds in Croatia from October 2005 until March 2006 as well as genetically analyzed and compared representative virus isolates to better understand the spread and epidemiology of this infection. This information may be useful for prevention and control of HPAIV H5N1 infection elsewhere.
Materials and Methods
Samples
A total of 6142 wild bird and 844 poultry samples were collected from September 2005 until June 2006. Most of the wild bird samples were examined for AIV within the frame of passive surveillance across Croatia. Remaining samples from wild birds and poultry were taken within active surveillance in the areas where HPAIV subtype H5N1 had been previously confirmed. Mainly organs (brain, trachea, lungs, liver, spleen, kidneys, and intestines) and sporadically cloacal and tracheal swabs were tested. Live birds and poultry were tested exclusively by tracheal and cloacal or, in some cases, only by cloacal swabbing.
Virus isolation
Virus isolation in chicken embryos and virus isolate subtyping by hemagglutination inhibition and neuraminidase inhibition tests were carried out according to the standard procedures (OIE 2004, Terregino and Capua 2009). A panel of monospecific antisera covering all 16 H subtypes and 9 N subtypes of AIV (Istituto Zooprofilattico Sperimentale delle Venezie, Padova, Italy) and monospecific sera against avian paramyxoviruses (APMV-1, -2, -3, -4, -6, -7, -8, and -9; Veterinary Laboratories Agency, Weybridge, UK) were used.
Molecular methods
Viral RNA for selected H5 isolates was extracted from allantoic fluids using High Pure Viral Nucleic Acid Kit (Roche Applied Science, Mannheim, Germany). Two-step reverse transcription and polymerase chain reaction (PCR) were carried out using GeneAmp® Gold RNA PCR Core Kit (Applied Biosystems Foster City, CA) according to the manufacturer's instructions. cDNAs were synthesized using random hexamers. Three partially overlapping fragments spanning the whole HA gene were amplified for each isolate by PCR using three primer pairs (Table 1), each in a final concentration of 1 μM, and the thermocycling conditions consisted of initial denaturation at 95°C for 5 min, followed by 40 cycles of 94°C for 30 s, 56°C for 30 s, 72°C for 3 min, and a final extension at 72°C for 10 min. All PCR products were electrophoresed in 2% agarose gel with addition of ethidium bromide and visualized on UV transilluminator. Specific bands were excised and purified with Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI). Sequencing of all PCR amplicons was performed in both directions with Big Dye Terminator v3.1 Sequencing Standard Kit (Applied Biosystems) in a 3730XL DNA sequencer (Applied Biosystems) and with same primers used for PCR. After sequencing, the primer sequences were trimmed off. The trimmed sequences for each virus isolate were assembled with ALIGN Plus 2.0 software. Phylogenetic analysis was performed with MEGA software version 4 (Tamura et al. 2007) by using Neighbor Joining algorithm with 1000 bootstrap replicates.
Based on GenBank accession number EF205157 (A/goose/Krasnoozerskoe/627/05 [H5N1]).
Number of noncomplementary nucleotides preceding or exceeding the hemagglutinin gene complementary sequences.
Nucleotide sequences were also translated into amino acid sequences to predict pathogenicity of each analyzed isolate according to the HA0 cleavage site.
Results
During the investigation period, 17 AIV subtype H5N1 were isolated from wild birds on six sites in four major areas. No AIV was isolated from Croatian poultry. Details about location, date, bird species, bird health status, and type of surveillance regarding virus isolation are shown in Table 2. Results of phylogenetic analysis of selected isolates from all four areas and all affected bird species are shown in Figures 1 and 2. All analyzed AIV subtype H5N1 isolates from Croatia during 2005–2006 possessed multiple basic amino acids at the HA0 protein cleavage site (PQGERRRKKR*GLF), indicating high pathogenicity of the avian influenza isolates (Rott 1992).

Phylogenetic tree of the hemagglutinin (HA) gene (nt 32–735) of highly pathogenic avian influenza virus (HPAIV) (H5N1) clade 2.2 (WHO/OIE/FAO H5N1 Evolution Working Group 2008). The tree was constructed using Neighbor Joining algorithm with 1000 bootstrap replicates. Length of the horizontal lines is proportional to the genetic distance among isolates. Scale bar indicates substitutions per site. Isolates from Croatia are indicated in bold and index case isolate is underlined. The text beside Croatian isolates describes date and place of virus isolation. GenBank accession numbers for index case isolate and isolates from other countries are given in parentheses. The tree is rooted with the A/bar-headed goose/Qinghai/0510/05 (H5N1) HA sequence (nt 32–1707) shown as an outgroup.
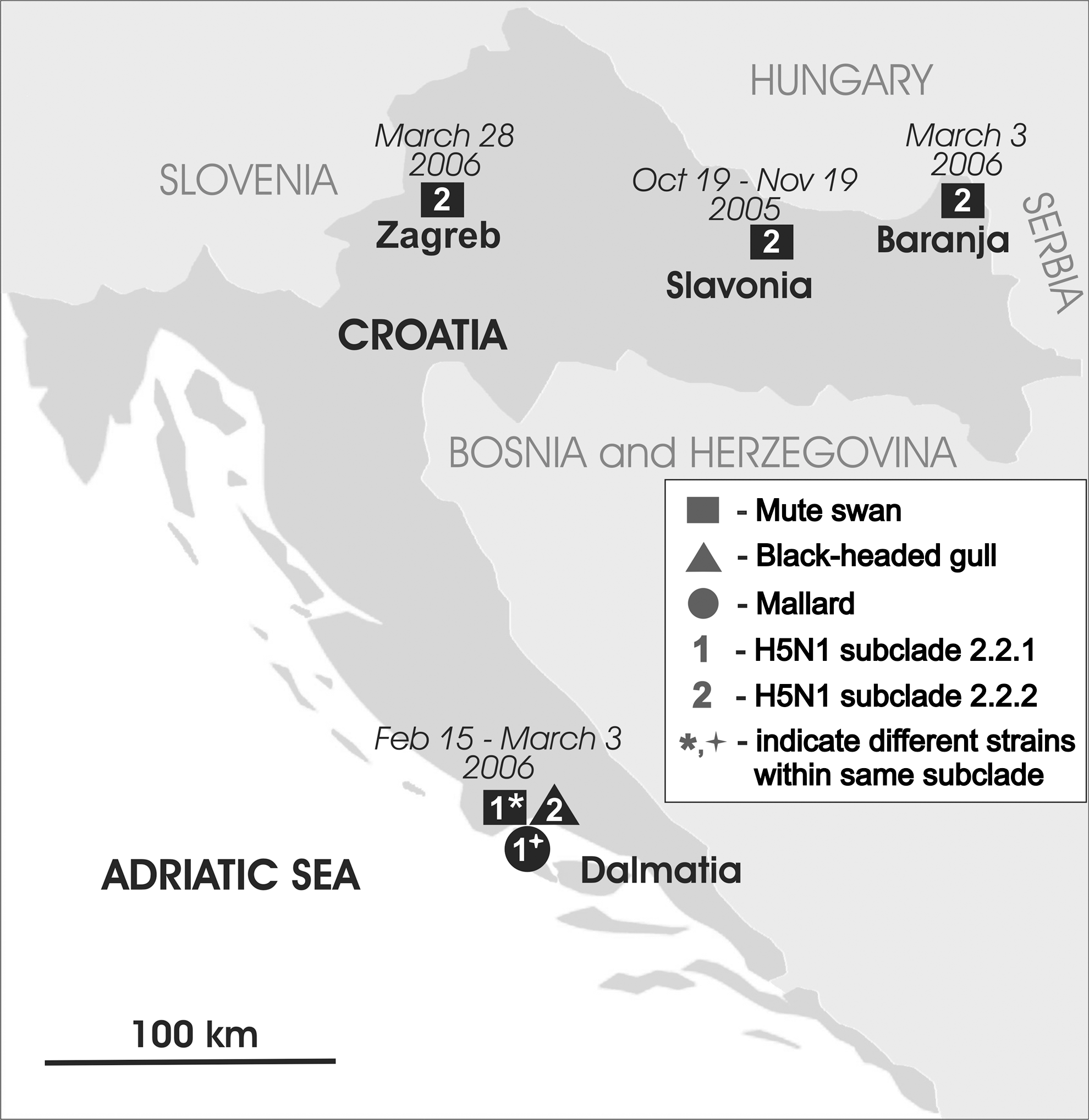
Map of Croatia with locations and time of outbreaks, affected bird species, and genetic subclades of analyzed highly pathogenic avian influenza virus (HPAIV) (H5N1) isolates. Squares, circle, and triangle represent mute swans (Cygnus olor), mallard (Anas platyrhynchos), and black-headed gulls (Larus ridibundus), respectively. Numbers 1 and 2 represent subclade 2.2.1 isolates and subclade 2.2.2 isolates, respectively (WHO/OIE/FAO H5N1 Evolution Working Group 2008), while asterisk and cross denote different genetic strains within the same subclade (2.2.1), suggesting at least three different introductions of H5N1 into Croatia.
Died with symptoms: sick bird was noted and tested after it died; Carcass: dead bird tested without known health history; Healthy: tested bird did not show any disease symptoms at the moment of sampling.
Individual samples were processed as a pool.
Isolates of highly pathogenic avian influenza virus that were genetically analyzed.
Pharingeal swab was also taken, but gave negative result.
The samples from the Pantana marsh were taken within active surveillance after positive finding in mute swan in the nearby village of Slatina.
In addition to AIV subtype H5N1, a low pathogenic AIV subtype H5N3 with HA protein cleavage site motif PQRETR*GLF and AIV subtype H10N7 were isolated from mallards (Anas platyrhinchos), whereas one PMV-1 was isolated from a mute swan.
Discussion
HPAIV H5N1 was confirmed in dead mute swans in Croatia on fish ponds “Grudnjak” and “Ribnjak 1905” in Slavonia in October 2005, on the island of Čiovo and nearby marsh of Pantana in Dalmatia in February 2006, as well as in Baranja and Zagreb in March 2006. HPAIV H5N1 virus was also confirmed in apparently healthy black-headed gulls (Larus ridibundus) in the marsh of Pantana at the end of February and early March 2006 as well as in a dead mallard from the same location in early March 2006. As anticipated, all seven analyzed Croatian isolates are H5N1 Qinghai-like viruses, that is, they belong to clade 2.2 (WHO/OIE/FAO H5N1 Evolution Working Group 2008). Most of these isolates belong to subclade 2.2.2 and are closely grouped with the index case isolate in Croatia (A/Cygnus olor/Croatia/1/05) from dead swans found on the fish pond “Grudnjak” (Fig. 1). Other members of this group are isolates from dead swans from Baranja and Zagreb and the isolate from apparently healthy gulls from the marsh of Pantana. Interestingly, five out of six H5N1 isolates from the reemerged outbreak in wild birds at the Qinghai Lake in China in May 2006 (QH06) were adjacent to A/Cygnus olor/Croatia/1/05, whereas one was related to the strain A/duck/Novosibirsk/02/05 from Russia (Wang et al. 2008). Finding that A/Cygnus olor/Croatia/1/05-like viruses were dominant H5N1 viruses circulating in Croatia during 2006 corroborates that the QH06 viruses of the Qinghai Lake may travel back via migratory birds (Wang et al. 2008). Other three analyzed isolates from Croatia belong to subclade 2.2.1 but, unlike Croatian isolates from subclade 2.2.2, which circulated in Croatia apparently for 6 months and remained genetically close, subclade 2.2.1 isolates show greater diversity and indicate that there are two distinct genetic strains (Fig. 1). This diversity of concurrent isolates is a result of independent evolution of the viruses that most likely took place before introduction into Croatia. Therefore, phylogenetic analysis of Croatian isolates from October 2005 until March 2006 implicates at least three independent introductions of H5N1 viruses by wild birds. Similarly, multiple introductions of H5N1 Qinghai-like viruses occurred in Germany (Starick et al. 2008) and in France (Gall-Reculé et al. 2008) in early 2006, but with poultry infections involved in the outbreaks.
The dominant A/Cygnus olor/Croatia/1/05-type strain of H5N1 virus in Croatia was found in all four affected areas (Slavonia, Baranja, Dalmatia, and Zagreb) and was the only detected strain in Slavonia, Baranja, and Zagreb. However, all three types were found in three different bird species near the town of Trogir in Dalmatia (Fig. 2). The first finding of H5N1 in Dalmatia was on 15 February, 2006 in a swan carcass on the seashore near Slatina village on the island of Čiovo. The carcass was obviously washed by sea stream from the neighboring marsh of Pantana about a kilometer away. A week later another swan carcass was found in the marsh of Pantana and the virus was phylogenetically similar to the virus isolated the previous week, indicating that both isolates were epidemiologically linked. Nevertheless, both isolates were significantly different from other H5N1 viruses isolated in Croatia. Subsequent active surveillance in the marsh of Pantana resulted in isolation of H5N1 viruses from apparently healthy black-headed gulls and from a dead mallard. The gulls were captured by mist nets and were released immediately after sampling and ringing (ring numbers available on request). Isolates from gulls were of A/Cygnus olor/Croatia/1/05-type strain, whereas the isolate from the mallard differed from all other H5N1 isolates from Croatia and was close to certain isolates from Western Europe. Finding of three different genetic strains of H5N1 virus at the same time in the marsh of Pantana, which is located on the Adriatic coast and has moderate winters, could be associated with frozen water surfaces in the continental part of Croatia as well as in Eastern Europe in early 2006 and the movement of birds toward warmer areas. Assumption about unusual migration pattern of wild birds from the Black Sea basin in other parts of Europe due to harsh winter conditions during 2005–2006 was also mentioned by Kilpatrick et al. (2006) and Alexander and Capua (2008).
During 2005–2006, three genetic strains of H5N1 virus were found in three different wild bird species; however, no particular relationship between the virus strain and the bird species was found, indicating cross-species transmission of the virus. Isolation of H5N1 virus from black-headed gulls is not common, especially from apparently healthy wild birds. Most of H5N1 virus detections were from dead or severely ill birds, including documented isolations of H5N1 virus from black-headed gulls (Ellis et al. 2004). Therefore, this is the first documented isolation of Asian HPAIV of H5N1 subtype from apparently healthy black-headed gulls. It is possible that the gulls were captured in the incubation phase while they shed the virus before exhibiting clinical signs of the infection. It is not known whether the gulls would have shown clinical sign or died later because they were released immediately after sampling and were not found afterward. If black-headed gulls do not exhibit clinical signs after infection with H5N1 virus and if they shed the virus, they is a possible link for virus transmission among wild birds. Pathogenicity of Asian HPAIV of H5N1 subtype for certain gull species apparently increased through viral evolution between 1997 and 2005 from asymptomatic or mild infection to fatal disease (Perkins and Swayne 2003, Brown et al. 2008a, 2008b). This implicates an assumption that the black-headed gulls captured in the marsh of Pantana were in the incubation phase and died afterward. Experimental infection of black-headed gulls with appropriate virus could give a reliable answer to this question.
Footnotes
Acknowledgments
All activities regarding sample collection, AIV detection, and field epidemiological investigation were funded by the Ministry of Agriculture, Fisheries, and Rural Development of the Republic of Croatia. Genetic analysis of the AIVs was funded by the Ministry of Sciences, Education, and Sports of the Republic of Croatia within the research grant “Genetic characterization of AI and ND viruses isolated in Croatia” (048-0481153-1136).
Disclosure Statement
No competing financial interests exist.