
Abstract
Five hantaviruses are known to circulate among rodents in Europe, and at least two among insectivores. Four (Dobrava, Saaremaa, Seoul, and Puumala [PUUV] viruses) are clearly associated with hemorrhagic fever with renal syndrome (HFRS). PUUV, the most common etiological agent of HFRS in Europe, is carried by the bank vole (Myodes glareolus), one of the most widespread and abundant mammal species in Europe. This host–virus system is among hantaviruses also the most studied one in Europe. However, HFRS incidence varies throughout the continent. The spatial as well as temporal variation in the occurrence of HFRS is linked to geographic differences in the population dynamics of the reservoir rodents in different biomes of Europe. While rodent abundance may follow mast seeding events in many parts of temperate Europe, in northern (N) Europe multiannual cycles in population density exist as the result of the interaction between rodent populations and specialist predator populations in a delayed density-dependent manner. The spatial distribution of hantaviruses further depends on parameters such as forest patch size and connectivity of the most suitable rodent habitats, and the conditions for the survival of the virus outside the host, as well as historical distribution patterns (phylogeographies) of hosts and viruses. In multiannually fluctuating populations of rodents, with population increases of great amplitude, one should expect a simultaneous build-up of recently hantavirus-infected (shedding) rodents. The increasing number of infectious, virus-shedding rodents leads to a rapid transmission of hantavirus across the rodent population, and to humans. Our review discusses these aspects for PUUV, the only European hantavirus for which there is a reasonable, yet still far from complete, ecological continental-wide understanding. We discuss how this information could translate to other European hantavirus–host systems, and where the most important questions lie for further research.
Introduction
SEOV, Seoul virus; DOBV, Dobrava virus; SAAV, Saaremaa virus; PUUV, Puumala virus; TULV, Tula virus; HFRS, hemorrhagic fever with renal syndrome.
Biology and ecology of the Puumala virus (PUUV)–bank vole system have been analyzed in numerous studies. For the other European hantaviruses this is much less, and published information is mostly limited to clinical or epidemiological descriptions of human cases or reports following rodent investigations not specifically aimed toward these other hantaviruses. Given the sometimes very different biology of the host rodents, such as the species' habitat requirements, population dynamics, and role in ecosystems, it is not all clear whether our knowledge about PUUV as such can be extended to the other hantaviruses as well. Data on Saaremaa virus (SAAV) and Tula virus are scanty, but for Dobrava virus (DOBV) we can try to discuss some patterns.
Hantaviruses and Their Hosts in Europe
Seoul virus (SEOV) infections appear very rare in Europe (Table 2) and there are no reported human clinical HFRS cases associated with SEOV, apart from a number of laboratory infections acquired from laboratory rats more than 20 years ago (Desmyter et al. 1983, Heyman et al. 2002b, Vapalahti et al. 2003). However, one antibody-positive human has been identified in France (Lundkvist, pers. comm.), and SEOV antibodies have been detected in Rattus rattus and R. norvegicus in Portugal (for references see Table 3) and in Belgium also SEOV viral genome in R. norvegicus (Heyman et al. 2009a).
Modified from Vapalahti et al. (2003), Kallio-Kokko et al. (2006), Bi et al. (2008), and Heyman and Vaheri (2008).
Cases/year = average no. HFRS cases per year of official reporting.
Cases/year = average no. HFRS cases per year of official reporting.
Cases peak year = no. HFRS cases peak year.
Peak year = year of highest record of HFRS.
Seroprevalence (%) = seroprevalence in human populations as reported per country or region, see footnotes below.
In foresters in Île-de-France.
Hantaan-like virus infection, with a rate of 2.3% in a healthy population.
In patients without symptoms of NE in Hattfjelldal, an endemic area.
Only one serologically confirmed case reported.
Only one HFRS case possibly related to TULV reported in Switzerland.
0% in forestry workers, blood donors 0.5%, farmers 0.8%, hunters 1.1%, and soldiers 1.9%.
DOBV is the hantavirus associated with the most severe form of HFRS in Europe, with a case fatality rate of about 10%. Nevertheless, human cases are rare and appear to be restricted to south-eastern (SE) Europe within only a part of the range of A. flavicollis (Plyusnin et al. 2006) (Table 2, Fig. 2a; see also section below).
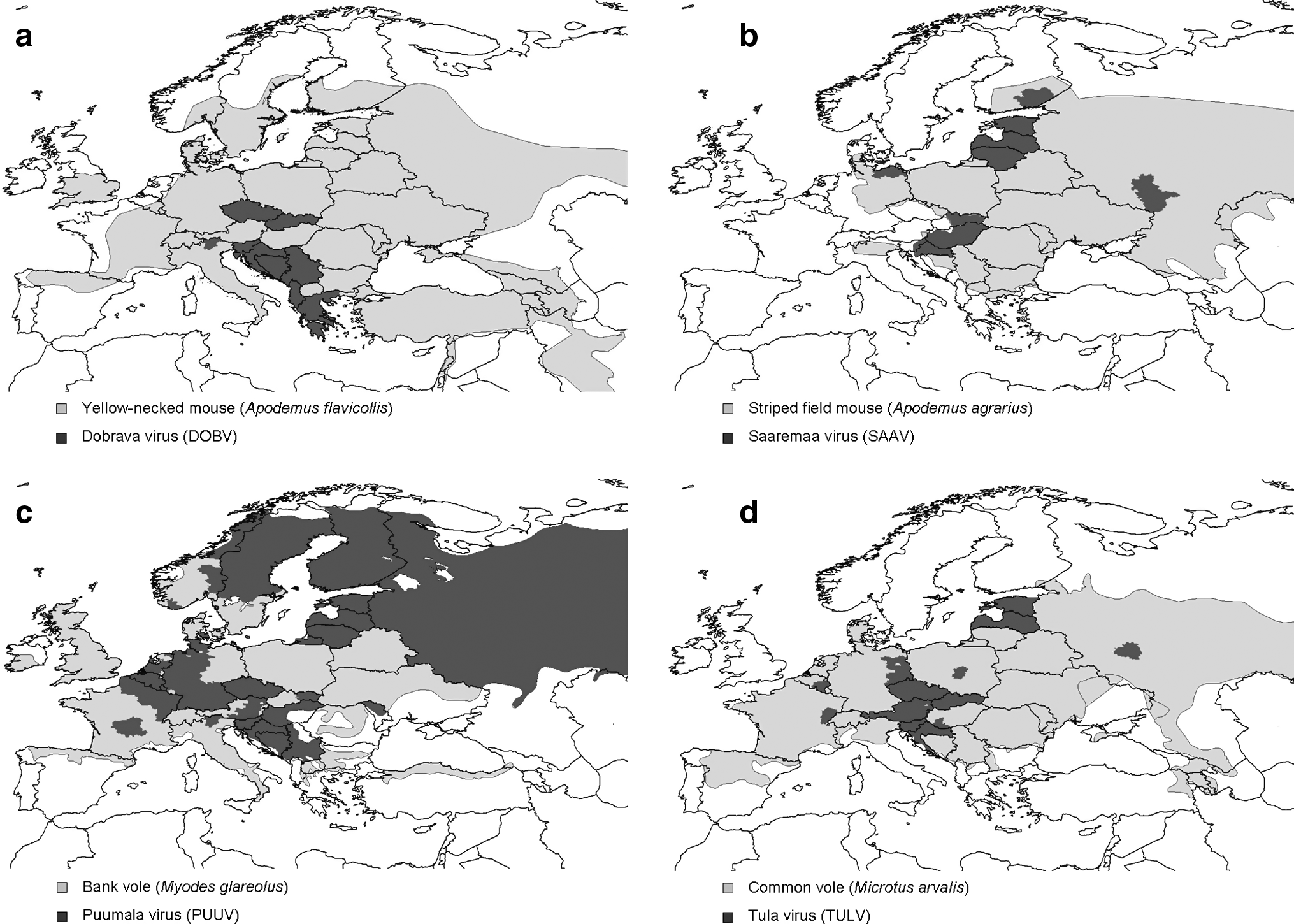
Geographical distribution of hantaviruses found in Europe and distributions of their reservoir hosts. Light gray indicates rodent host species present; dark gray indicates virus present or origin of hemorrhagic fever with renal syndrome cases; no fill indicates no data or neither host or virus present. (
While most human SAAV infections have been found with serological studies in Estonia (for references see Table 3), viral genome has also been found in rodent samples from Estonia, Germany and Russia, Denmark, Hungary, Slovenia, Slovakia (for refs see Table 3), and Croatia (Markotić, Henttonen, and Plysunin, pers. comm.) and one seropositive specimen in Southeastern Finland (Fig. 2b). Clinical HFRS caused by SAAV is rare, but outbreaks have occurred in European Russia (Plyusnin et al. 1999, at the time assumed to be DOBV infections), and the regionally relatively high SAAV seroprevalence among humans suggests a usually subclinical or mild course of infection and/or unawareness of the clinical signs among local physicians (Brus Sjölander et al. 2002, Golovljova et al. 2002) (Table 2), and this may also apply to, for example, the SEOV system. A debate on DOBV/SAAV phylogeny has occurred, and more geographic lineages need to be studied. The original SAAV is found in A. agrarius on some islands in the Baltic; the strain in A. agrarius in mainland Europe is often included in SAAV, sometimes called “DOBV aa”; and a third strain, DOBV or “DOBV af,” is found in A. flavicollis in SE Europe.
PUUV then is by far the most common etiological agent of HFRS in Europe. In Northern Europe thousands of PUUV–HFRS cases (commonly referred to as nephropathia epidemica) are reported every year (Brummer-Korvenkontio et al. 1980, 1982, 1999, Vapalahti et al. 2003) (Tables 2 and 3), and an apparent increase in incidence has occurred over the past decade, partly due to increased public and clinical awareness also in regions where the disease was not recognized before, in parallel with the development of better diagnostic tools (Olsson et al. 2007, 2009, Heyman and Vaheri 2008, Pettersson et al. 2008, Vaheri et al. 2008). However, it needs to be investigated whether these factors explain the observed upsurge alone; in Finland a change in geographic synchrony of bank vole population cycles, from previously more localized cycles to an almost nationwide synchrony today, has probably contributed to the observed pattern (Henttonen and Vaheri 2006). Recent PUUV-HFRS outbreaks in Western Europe (Heyman et al. 2007) are presumed to be caused from a causal connection of higher summer temperatures leading to increased frequency of masting of European beech (Fagus sylvatica) and pedunculate oak (Quercus robur), providing an abundance of food for bank voles allowing for population increases and supporting PUUV transmission (Tersago et al. 2009, 2010) (see also section below).
Tula virus is a fifth hantavirus found in many parts of Europe (Song et al. 2004). It is associated with the common vole (Microtus arvalis; Fig. 2d) and with other sympatric Microtus species (Plyusnin et al. 1994, Plyusnina et al. 2007, 2008, Schmidt-Chanasit et al. 2010), and human infections without clinical disease have been documented by serological tests (Heyman et al. 2002a, Klempa et al. 2003, Vapalahti et al. 2003, Vaheri et al. 2008).
The role of insectivore (order Soricimorpha) species as hantavirus reservoirs has only begun to be explored. Song et al. (2007) established the connection of Seewis virus to the Eurasian common shrew (Sorex araneus), and Kang et al. (2009) did similarly to Nova virus and the European common mole (Talpa europaea). Both these insectivore species are widely distributed on the European continent, but it remains to demonstrate if they cause disease to humans.
Geographical Distribution
PUUV is found in large parts of Northern Europe: Finland (for references, see Table 3), Sweden and Norway, parts of European Russia, the Baltic States Estonia, Lithuania, and Latvia. In remaining Europe PUUV has a patchy distribution within, for example, a region constituted by Belgium, Luxembourg, and eastern and southern parts of the Netherlands, northeastern France, and the Central Massive, as well as western/southern Germany. The reservoir of PUUV, the bank vole, is one of the most widespread and abundant mammal species in Europe and is found in most areas except the northernmost regions and near the coasts of the Mediterranean Basin (Fig. 2c, modified from Mitchell-Jones et al. (1999), Temple and Terry (2007), Kotlik et al. (2006)). Indigenous cases of nephropathia epidemica and/or PUUV-infected rodents are also found in Denmark, the Czech Republic, Slovakia, Austria, Hungary, northern Italy, and in Southeastern Europe, for example, Croatia, Bosnia-Herzegovina, Serbia and Montenegro, Slovenia, and Greece (Table 2, Fig. 2c). There are also PUUV-specific antibodies detected from retrospective analysis in patients with hypertransaminemia in Spain.
Following the latest glacial period, the bank vole (and other terrestrial mammals) recolonized the continent through various routes, giving rise to well-studied phylogeographic patterns (Asikainen et al. 2000, Deffontaine et al. 2005). These patterns of a northern recolonization of bank voles from Ural regions via present Finland to northern Fennsocandia (here northern parts of Finland, Sweden, and Norway) and southeastern recolonization up to central Finland, and, respectively, a southern route over the at the time present land corridor between present Denmark and Sweden are also apparent in the phylogeography of PUUV (Plyusnin et al. 1995, Hörling et al. 1996, Sironen et al. 2002, Johansson et al. 2008, Razzauti et al. 2009). However, the present-day PUUV and bank vole mt-DNA lineages are mostly not congruent, except for the suture zone in north-central Sweden (Razzauti et al. 2009, Nemirov et al. 2010). This suggests that there may have been switches between lineages during the multiglacial history of N Europe.
It remains an enigma why PUUV and many other hantaviruses seem to be restricted to a limited portion of the host species' distribution (Fig. 2a–d). However, studies on phylogeny and postglacial recolonization patterns of bank voles (Tegelström 1987, Nemirov et al. 2010) and PUUV (Hörling et al. 1996, Nemirov et al. 2010) clearly suggest that there was a past presence of PUUV in, for example, southernmost Sweden in bank voles of Western lineage. Neither confirmed autochthonous HFRS cases nor PUUV in bank voles (Olsson, pers. obs.) has been reported from southernmost Sweden, but PUUV of South Scandinavian lineage was found further north in bank voles of Western lineage (Nemirov et al. 2010).
The distribution of DOBV is seemingly also limited to just a part of its host's, the yellow-necked mouse, range (Fig. 2a) and the underlying mechanism remains unclear. The perceived absence or low prevalence of DOBV in most parts of Europe (Kallio-Kokko et al. 2006) reveals a need for more comprehensive investigations on the topic (but see Papa and Antoniadis 2001). However, according to observations by Michaux et al. (2004), the yellow-necked mouse went extinct during the last Quaternary glaciations in Western, Eastern, and Northern Europe, and the only prevailing populations survived in refuges of the Italo-Balkan region (in three genetic subclades), and in the Near and Middle East. In the following postglaciation era, the whole Western Palearctic area was recolonized from the Italo-Balkan populations. Such expansions from refugial populations also involve serial bottlenecks, and one could speculate that, parallel to the observed loss of genetic diversity among yellow-necked mouse populations living in the more recently colonized regions (Michaux et al. 2004), also specific horizontally transmitted viral pathogens such as DOBV may be lost during these bottleneck events. Still, the question remains why DOBV then has not spread again from the areas where it is currently present. One hypothesis is that different hantaviruses are unequally tolerant to occurring ecological conditions. An alternative explanation could be that host densities are generally too low, or populations too fragmented to sustain the infection permanently. There remains also the option that the virus is present but simply has not yet been detected.
Regional and Temporal Host Population Dynamics
There is a strong positive correlation between bank vole numbers and the risk of humans acquiring PUUV-HFRS (nephropathia epidemica). In Finland and northern Scandinavia, bank vole populations show multiannual cyclic patterns of 3–4 years with increase, peak, and decline/low phase (Henttonen et al. 1985, Olsson et al. 2003a, Hörnfeldt 2004, Korpimäki et al. 2005). In Sweden, for example, bank vole abundance alone explained >70% of the variation in seasonal HFRS incidence (Olsson et al. 2009), and in Finland the current increase or peak phase of the bank vole cycle was a good predictor of risk (Kallio et al. 2009). During rapid population growth, either in cyclic increase phase or due to masting, one should expect a simultaneous build-up of recently hantavirus-infected and -shedding rodents that cause a rapid frequency-dependent transmission of hantavirus across the rodent population, and subsequently increased risk to humans (Davis et al. 2005; Sauvage et al. 2006).
The seasonal pattern of PUUV infection among bank voles appears similar in N Europe and temperate Europe in nonoutbreak years, that is, highest PUUV antibody prevalence in spring and among adult individuals, and since winter breeding among bank voles is rare, the spring populations consists almost exclusively of over-wintered adult voles (Escutenaire et al. 2002, Olsson et al. 2002, 2003a, Kallio et al. 2007, 2009, Augot et al. 2008, Razzauti et al. 2008), but, importantly, this pattern is seasonal, not density dependent. Still, at high densities the absolute numbers of seropositive voles peak. In temperate zone during mast-induced outbreaks, bank voles breed late, even in winter, and early next spring, and hence increased seroprevalence due to high breeding activity occurs already in autumn and winter (Tersago et al. 2010). It is important to understand that different ecological factors affect bank vole population dynamics in mainland temperate Europe as compared to boreal northern Europe; therefore, it is relevant to emphasize the biome-specific host and virus dynamics (see sections below). Although changes in rodent dynamics may be occurring (Ims et al. 2008), vole population dynamics appear to be controlled by top-down processes in N Europe (i.e., controlled by interactions with predators) versus bottom-up processes (i.e., controlled by food availability) in many other parts of Europe (Hansson et al. 2000, Hanski et al. 2001, Lambin et al. 2006).
There are sparse data available concerning seasonality and interannual variation of HFRS caused by the other hantaviruses in Europe. However, observations on DOBV infections, in SE Europe, reveal that most cases occur during summer months June to August (coincident with, e.g., crop harvesting), and noticeably higher numbers of HFRS cases in 11 years in the 48-year period from 1961 through 2008 (Avšič-Županc 1999, Heyman et al. 2009b, Avšič-Županc, pers. comm.). There appears to be a periodicity that may be influenced by, for example, masting events. Although some ecological characteristics of the yellow-necked mouse in Europe are distinct from the bank vole (i.e., it is mainly granivorous and geographically confined to temperate broadleaf and mixed forests), its population dynamics (e.g., increases in population density following mast seeding) are similar (Hansson 1985, Pucek et al. 1993, Bujalska and Grüm 2008, Tagliapietra et al. 2009).
Northern Europe
Throughout the extensive boreal taiga of northern Fennoscandia population, dynamics of bank voles and other sympatric voles are synchronized by specialist predators, causing a 3–4-year cycle with population peaks and troughs of both voles and, with delay, these predators. Here, north of 60–61° N, snow-cover may persist >120 days/year. The climate shapes the structure of mammal communities, leading to limited diversity prey guilds (mainly voles and also lemmings in north) preyed upon by specialist predators. This interaction results in time lags in the population dynamics of prey and predators (Hansson and Henttonen 1985, Hanski et al. 1991, 2001, Ims and Fuglei 2005, Kausrud et al. 2008) with peak prey densities usually occurring every 3 or 4 years (Henttonen et al. 1985, Olsson et al. 2003a, Hörnfeldt 2004, Korpimäki et al. 2005). As contrast, in temperate zone like Southern (S) Scandinavia and many parts of mainland Europe, a higher diversity in community structure of prey and predator guilds promotes population stability. The snow-cover also provides seasonal refuge for voles from several predators (Hansson and Henttonen 1985, 1988, Hanski et al. 1991, 1993, 2001, Henttonen 1991, Hanski and Henttonen 2002, Korslund and Steen 2006), and probably also from harsh weather.
It has been long recognized that infestation of human-made peri-domestic refuges by voles in late autumn–early winter plays a critical role in the vole to human transmission of PUUV (Nyström 1977, Korpela and Lähdevirta 1978, Vapalahti et al. 2010). A majority of HFRS patients who can identify the time and place of exposure to PUUV reported this exposure to have occurred in or near a dwelling, often in association with activities where they came in contact with dirt and dust contaminated by rodent excreta, such as handling of fire wood, redecorating, or cleaning (Olsson et al. 2003b). However, winters in the north have become warmer (Alexandersson 2002, Kausrud et al. 2008), particularly with sudden warm periods when the snow-cover melts and subsequent freezing causing voles to migrate and infest human dwellings also later in winter (Olsson et al. 2007, 2009).
In the north there are also differences in habitat use by bank voles and probability of encountering PUUV-shedding bank voles during the periods when rodents normally do not utilize peri-domestic habitats. In particular, and to some degree similar to the observations in mainland Europe, the local abundance of bank voles and the number of PUUV-infected bank voles was highest in mature, moist forests characterized by conifers Picea abies (and connected vegetation like Alectoria spp. and Vaccinium myrtillus) and fallen wood (Olsson et al. 2005). Still, in the vole peak years bank voles and PUUV are found in all habitats from recent clear-cuttings to old forests (Voutilainen et al., pers. comm.).
Mainland Europe
Bank voles in temperate Europe show population irruptions following mast seeding events and subsequent increased abundance of food (Jensen 1982, Pucek et al. 1993, Tersago et al. 2009). Masting events are triggered by climatic factors that induce the formation of flower buds on, in particular, the European beech (Fagus sylvatica) and pedunculate oak (Quercus robur), and subsequent fruit setting and development of beech nuts and acorns. The climatic factors promoting masting in the European beech are high temperature combined with low precipitation in the summer (in particular July) during the year before masting (Övergaard et al. 2007), while heavy rains, frost, hailstorms, or strong wind may destroy the flower buds (Holmsgaard and Olsen 1960, Gruber 2003). Similarly, high temperature during summer of the previous year and during spring in the current year favors acorn production. Nevertheless, in contrast to its effect on beech nuts, summer drought during the previous year has a negative effect on acorn production. These contrasting effects of summer drought help explain why mast seeding not always occurs simultaneously for the two tree species (Gurnell 1993).
The frequency of beech masting appears to have increased over the past three decades (Övergaard et al. 2007). As a consequence of climate change and increased temperatures in the areas concerned, mast seeding events may continue to increase in frequency or may stabilize at this higher frequency (Övergaard et al. 2007). These conditions could support higher numbers of the rodents that serve as the principal hosts of PUUV and DOBV, and lead to a higher risk of HFRS in humans (Heyman et al. 2007). In fact, by analysis of the summer temperature during the previous years, beech masting can be predicted and thus related to food supply for rodents (Övergaard et al. 2007). Recently, Tersago et al. (2009) and Clement et al. (2009) showed that a rise in temperature during two consecutive summers is associated with higher HFRS incidence in Belgium and high bank vole numbers (Tersago et al. 2010). Such a predictive risk tool is likely to be applicable also in other geographic areas where mast seeding events are coupled to an increase in rodent numbers and HFRS.
Environmental Conditions
Environmental factors also influence the local PUUV persistence on a finer spatial scale. The prevalence of PUUV infection was higher among bank voles in preferred environments, such as wet or very humid habitats compared to dryer habitats within the same study area (Verhagen et al. 1986, Linard et al. 2007b), where the virus occurrence was related to soil characteristics while host occurrence was related to landscape fragmentation.
The general educated guess concerning survival of hantaviruses outside the host has been approximately 2 days, based on hantaviruses being lipid-enveloped single-strand RNA viruses (Schmaljohn et al. 1999), but this assumption was disputed by theoretical modeling (Sauvage et al. 2003). The length of viral infectiousness was evaluated in controlled experiments where naïve bank voles were exposed to bedding material from cages of PUUV-infected specimens, and it was found that bedding material contained infectious PUUV for 12–15 days at room temperature (Kallio et al. 2006a). Using additional cell culture experiments, Kallio et al. (2006a) showed that low temperature and high moisture promoted virus longevity. Therefore, during autumn–winter conditions in Northern Europe, PUUV may remain infectious outside the host for several weeks. A subsequent experiment on the temporal patterns and quantitative levels of PUUV shedding, for example, RNA in saliva, urine, and fecal excretions of experimentally inoculated naïve bank voles, showed that PUUV RNA levels peaked between the 2nd and 4th week postinfection (pi) but remained at detectable levels in 84, 44, and 44 days pi in saliva, urine, and feces, whereas PUUV RNA was found in the blood 133 days pi (Hardestam et al. 2008). The relevance of these laboratory findings must be further investigated under field conditions.
Host and Viral Genetics
The majority of field studies on bank voles have focused on the influence of extrinsic factors such as climate and habitat on PUUV occurrence at the population level (e.g., Olsson et al. 2005, Linard et al. 2007a, Tersago et al. 2008), and on the effects of host dynamics on PUUV infections in bank voles and in humans (those above, Brummer-Korvenkontio et al. 1982, Kallio et al. 2009). Recent studies, however, suggest that the rodent host susceptibility to PUUV infection may depend also on intrinsic individual factors such as genetic background (Deter et al. 2008a) and kinship among bank voles (Deter et al. 2008b). The susceptibility of a bank vole to PUUV infection may depend on which major histocompatability complex (MHC) allele it carries (Deter et al. 2008a). In addition, the kinship between PUUV antibody-positive bank voles was higher than that between antibody-negative animals (Deter et al. 2008b). These observations on genetics and kinship, however, need be clarified and validated by laboratory studies. These observations suggest that not all bank vole genotypes are equally susceptible to PUUV infection and that the persistence of PUUV in local populations may be influenced by population genetics as well as extrinsic factors. Thus, the confinement of PUUV within a restricted refugium may not only be due to prime habitat composition but also to the prevalence of susceptible bank vole immunogenetics.
A better understanding of the phylogeography of hantaviruses and their hosts will lead to a clearer understanding of their evolutionary and dispersal history (Plyusnin et al. 1995, Sironen et al. 2001, Razzauti et al. 2009, Nemirov et al. 2010). PUUV genetics may show substantial variation also within highly endemic areas and short distances. Within a region of very high HFRS incidence (313/100 000 in 2007, Olsson et al. 2009) PUUV genome sequences appeared stable for >80 km in two different phylogenetic clusters on either side of an apparent contact zone, but differed by 7% in the viral S-segment between two sampling sites separated by only 10 km on either side of this contact zone (Johansson et al. 2008).
Effects of Hantavirus Infection on Host Life History
Hantavirus infection is usually assumed to be nonpathogenic in the natural host. Recent work, however, showed that PUUV-infected bank voles had a significantly lower overwinter survival probability than antibody-negative bank voles (Kallio et al. 2007). This could obviously affect bank vole population dynamics and demography, which in turn again would affect transmission patterns. The role of such complex interactions between infection and host population need to be further investigated, and special emphasis should be addressed the impacts of infection in different seasons and population subgroups (functional groups).
PUUV-induced maternal antibodies (MatAb) may also play an important role on transmission patterns. While it has been shown that presence of MatAb delay the horizontal infection of PUUV in bank voles until the approximate age of 3 months (Bernshtein et al. 1999), and that MatAb's really delay transmission process in wild bank vole populations (Kallio et al., in press), little was known about the effects of PUUV on the individual bank vole's life history traits and fitness. Recent studies have shown that bank voles with PUUV MatAb's became sexually mature earlier and had a higher breeding success than voles without MatAb's (Kallio et al. 2006b). However, based on the results from island experiments where all or none of the founding females were PUUV infected, it was suggested that a large proportion of bank voles with MatAb may lead to local extinctions of PUUV as too few susceptible animals remain (Kallio 2006c). Much remains to be studied concerning (altered) behavior (intensified shedding), metabolic rate, and cost of the persistent elevation of an immune response among rodents infected by a hantavirus.
The Future
Within the pan-European project “Emerging Diseases in a changing European Environment” (EDEN,
The evident differences in incidence of human hantavirus infections by the various hantaviruses need be carefully investigated. Obviously, there is a virus-specific virulence to each hantavirus. However, there are likely also several other factors acting alone or in concert behind these patterns, such as differences in species abundance and behavioral ecology of specific hantavirus' host species. Bank voles, for instance, seasonally and regionally utilize peridomestic rodent harborage where other rodent species do not. This may give raise to a rodent species' specific direct or indirect contact rate with humans and subsequently also a difference in probability of human exposure to hantavirus in relation to the ecology of their hosts.
In conclusion, several studies report the assumed local absence of PUUV during periods of decreased bank vole abundance, while the virus seems to reestablish during population increases (e.g., Escutenaire et al. 2000, Olsson et al. 2003a, Kallio et al. 2009). One factor is thus not enough when determining the presence of PUUV, but the interplay of rate and amplitude by which the local bank vole population may increase locally seems to allow for a rapid increase in PUUV transmission between conspecifics during such population events, whether through direct or indirect transmission (Sauvage et al. 2006). The differences in landscape structure between boreal and temperate zones also affect the PUUV spread and extinction (Tersago et al. 2008). The virus transmission between conspecifics in relation to (1) frequency-dependent versus density-dependent processes, (2) direct versus indirect transmission, and (3) significance of the individual vole's demographic and genotypic traits on PUUV infection and the effects on virus persistence locally need to be further evaluated. Although the ecological conditions differ between the hantavirus and host systems, also among insectivores (order Soricimorpha), and in different biomes, the observations from studies of the PUUV-bank vole system are also useful for comparisons when investigating other hantavirus–host systems in Europe.
Footnotes
Acknowledgments
We want to express our gratitude to James N. Mills for commenting on earlier draft of this article, Mike Begon and an anonymous reviewers for their constructive comments. This work was partly supported by and partially funded by the Swedish Emergency Management Agency/Krisberedskapsmyndigheten (KBM), and EU grant GOCE-2003-010284 EDEN. The article is catalogued by the EDEN Steering Committee as EDEN 0149 (
Disclosure Statement
No competing financial interests exist.