
Abstract
Pigs have been confirmed to be reservoirs of some genotypes of hepatitis E virus (HEV), and other nonhuman species are also likely infected with the virus. To assess the prevalence of HEV infection in domestic animals in China, 3579 serum samples, including 1967 swine, 700 goat, and 912 cattle sera, were collected from 26 provinces across the country and tested for HEV antibodies and antigen using enzyme immunoassays. The results showed that 82.2% of the swine samples, but only 10.4% and 28.2% of cattle and goat sera, were anti-HEV positive respectively. The prevalence of anti-HEV antibody in animals varied from province to province, ranging from 10.9% to 100% in pigs, 0% to 48% in goats, and 0% to 92.9% in cattle. About 1.9% of pigs, 1.6% of goats, and 0.8% of cattle tested in the study were positive for HEV antigen. Some samples, including all HEV antigen–positive samples, were tested for HEV-specific RNA using reverse transcription polymerase chain reaction. Fifteen swine samples, but none from the goats or cattle, were found to be HEV RNA positive. Sequence and phylogenetic analyses classified all the swine HEV isolates into HEV genotype 4, which was further divided into four subgroups. This study demonstrated that HEV infection is widespread in domestic animals, particularly pigs, in China. The HEV genotype infecting pigs in China was genotype 4. However, the isolates displayed considerable genetic diversity.
Introduction
Natural antibodies against HEV have been detected in numerous animal species, suggesting the spread of HEV among these animals. Successful experimental transmission of human HEV to animals indicates that HEV can infect across species (Huang et al. 2008, Meng 2003, 2009). HEV sequences have been isolated from swine in many countries and, thus far, these isolates belong to genotype 3 or 4. Further, human infections with these two genotypes are found to be caused by HEV genetically closely related to, and in some cases identical to, genotypes 3 and 4 strains found in pigs in the same geographical area (Wang et al. 2002, Meng 2003). Accumulating evidence suggests that HEV infections in humans, at least those caused by genotypes 3 and 4, may be acquired zoonotically, and domestic pigs, and perhaps other animals, are reservoirs of the virus (Meng 2009).
Hepatitis E was first recognized in China after a large epidemic in the southern part of the Xinjiang Uighur autonomous region (Zhuang et al. 1991), and small epidemics and sporadic cases have been reported from various regions of the country. Thus, HEV is known to be endemic in China. The Xinjiang epidemic was attributed to HEV genotype 1, but genotype 4 has been isolated more often from sporadic cases in the country than is genotype 1 (Wang et al. 1999, Zhai et al. 2006, Yu et al. 2009, Tai et al. 2009). Reports suggest that HEV infection is endemic in domestic animals in some regions of China and the anti-HEV antibody is particularly highly prevalent in pigs. The aim of this study was to understand more fully the prevalence and genetic variety of HEV present in domestic animals in China, including pigs, goats, and cattle.
Materials and Methods
Serum samples
A total of 3579 serum samples, including 1967 swine, 700 goat, and 912 cattle sera, were collected from 26 provinces in China in the autumn of 2008. Most of the samples were collected from industrialized farms, but some were collected from small groups of animals raised by peasants. All the serum samples were stored in dry ice for transportation and then transferred to and stored in a −70°C freezer until use.
Detection of anti-HEV antibody and antigen
Anti-HEV antibody and antigen were detected by enzyme immunoassay (EIA), using a direct-sandwich EIA kit and a monoclonal antibody-–based EIA kit (Wantai Biopharmaceutical, Beijing, China), respectively, according to the manufacturer's instructions. Fifty microliters of each sample were used for antibody detection and 100 μL for antigen detection (Wang et al. 2002, Zhang et al. 2006).
RNA extraction
RNA was extracted from each sample using a spin-column kit and following the procedures of the Kinghawk Biopharmaceutical Company (Beijing, China). The protocols were essentially as described previously (Wang et al. 2002, Li et al. 2008), except that 100 μL serum were used for RNA extraction. The RNA from each sample was recovered in 50 μL of elution buffer.
Nested reverse transcription polymerase chain reaction
The nested reverse transcription polymerase chain reaction (RT-PCR) primers derived from the ORF2 region are capable of detecting divergent HEV strains (genotypes 1 to 4) and have been described previously (Li et al. 2008). The external primer pair was 5′-AAYTATGCWCAGTACCGGGTTG-3′ and 5′-CCCTTATCCTGCTGAGCATTCTC-3′; the internal primer pair was 5′-GTYATGYTYTGCATACATGGCT-3′ and 5′-AGCCGACGAAATYAATTCTGTC-3′. Nested RT-PCR was carried out in 50 μL reaction mixtures containing 7 μL RNA extract, 20 pmol of each of the external primers, 5 U AMV reverse transcriptase (from avian myeloblastosis virus), and 5 U Taq polymerase. The reaction was performed at 50°C for 30 min, followed by 35 cycles of amplification at 94°C for 1 min, 50°C for 45 s, and 72°C for 1 min, with a final extension at 72°C for 10 min. Two microliters of the PCR-amplified product were transferred into a second PCR reaction mixture with a total volume of 50 μL, containing 20 pmol of each of the internal primers and 5 U Taq polymerase. The amplification parameters were the same as for the first PCR. A final amplification product of 348 bp was then analyzed by electrophoresis through 1.0% agarose gel and viewed by ethidium bromide staining.
DNA sequencing and sequence analyses
Amplified PCR products were separated on 1.0% agarose gel. Bands of the predicted size were excised from the gel and purified using a Quick Spin™ column kit (Qiagen, Hilden, Germany). The purified products were sequenced directly on both strands with the specific PCR primers synthesized by Invitrogen (Beijing, China). Sequences were analyzed using ClustalW2 and Mega4 software. The swine HEV sequences were subsequently compared with selected reference sequences from human and swine HEV strains available in the GenBank database. These included the following: for genotype 1, Burma (M73218, human), China Hebei (M94177, human); genotype 2, Mexico (M74506, human); and genotype 3, US1 (AF060668, human), swJHK2-3 (AB194479, Japan, pig), SAAS-JDY5 (FJ527832, Shanghai, pig). The representative strains for each subgenotype of genotype 4 identified by Lu et al. (2006) were as follows: 4a, T21 (AF151963, China Beijing, human), Ch87 (AJ344171, China Beijing, human); 4b, Ch181 (AJ344188, China Beijing, human); 4c, swJ13-1 (AB097811, Japan, pig), swJ14 (AB080575, Japan, pig); 4d, Ch108 (AJ344181, China Beijing, human), swCH72 (AY596308, China Uighur, pig); 4f, HE-JA2 (AJ082558, Japan, human, pig); and 4g, CCC220 (AB108537, China Beijing, human). Chinese swine strains with complete genome sequences in GenBank were swCH189 (FJ610232, Gansu), swGX40 (EU676712, Guangxi), swGX32 (EU366959, China), SH-SW-2SI (EF570133, Shanghai), SW-DQ (DQ279091, Heilongjiang), swCH31 (DQ450072, Jiangsu), and swCH25 (AY594199, Jiangsu).
Statistical analysis
The chi-square test was used to compare HEV prevalence among provinces. To compare HEV antibody and antigen levels expressed as the S/CO (the ratio of Sample Value to Cutoff Value) mean ± standard error of mean (SEM), statistical analysis was performed using the unpaired Student's t-test. p-Values <0.05 were considered significant.
Results
Prevalence and levels of anti-HEV antibody in domestic animals
Of the 1967 swine sera collected from 25 provinces in China, 1617 (82.2%) were anti-HEV antibody positive (Table 1). Positive samples were found in all 25 provinces, and their prevalence varied considerably among the provinces, ranging from 10.9% to 100%. The lowest prevalence of 10.9% was for samples from Sichuan, and the neighboring province Chongqing also had a low prevalence (17.3%), both being significantly below the average.
Seven hundred goat serum samples collected from 13 provinces and 912 cattle serum samples collected from 17 provinces also were tested for anti-HEV antibody. The prevalence ranged from 0% to 48% in goats and 0% to 92.9% in cattle. The average prevalence for both species was significantly lower than for swine (Table 1, p < 0.001). However, some individual groups showed exceptionally high positivity rates. For example, 13 of 14 (92.9%) cattle sera from Guangdong were anti-HEV positive. The average titer of HEV antibody (expressed as S/CO mean ± SD) in pigs (10.7 ± 4.7) was also significantly higher than in goats (4.6 ± 2.9) and cattle (3.7 ± 3.4).
HEV antigen and HEV RNA in domestic animals
All serum samples were tested for the presence of HEV antigen. The positivity rate for pigs 1.9% (38/1967) was almost equal to that for goats 1.6% (11/700) but a little higher than for cattle 0.8% (7/912) (Table 1). Cattle sera from most provinces were negative except 6 of 495 samples from Inner Mongolia and 1 of 22 samples from Shanghai. The average titer of HEV antigen (expressed as S/CO mean ± SD) in pigs (3.94 ± 2.92) was statistically higher than in goats (1.85 ± 0.87) and cattle (1.90 ± 1.74).
On the basis of the report that HEV RNA is present mainly in HEV antigen–positive animal sera (Yu et al. 2009), in this study all of the HEV antigen–positive and HEV antibody–positive samples with OD values below 1.0 were tested for HEV RNA by RT-PCR. All goat samples from Shanxi and Guizhou, and all cattle samples from Guangdong, Xinjiang, Yunnan, and Tianjin with relatively high anti-HEV prevalence were also tested for HEV RNA. Fifteen swine sera were HEV RNA positive and all of them were also positive for HEV antigen. RNA positivity rate of the antigen-positive samples was 39.5% (15/38), and the overall prevalence of HEV RNA in pigs was 0.8% (15/1976). None of the goat or cattle samples was positive for HEV RNA.
Phylogenetic analysis of the HEV sequences
The PCR products from the 15 positive swine samples were sequenced. The 348 bp sequences of the ORF2 capsid region were compared and aligned with known HEV sequences and subjected to phylogenetic analysis. Sequence analysis showed that the 15 swine isolates had 82–100% nucleotide sequence identity with each other. All the 15 sequences clustered within genotype 4 and were further divided into four distinct subgroups as shown in the phylogenetic tree (Fig. 1). One isolate from Shanxi and one from Tianjin are in the 4a subgroup. Two identical isolates from Xinjiang and two from Shanxi cluster in the 4d subgroup. Three identical isolates from the same farm in Hangzhou and six from Fujian, Hubei, Guizhou, Tianjin, and Anhui cluster closely together and form a novel subgroup. An isolate from Jiangsu, along with other strains reported previously, falls into another subgroup. The nucleotide differences among the subgroups ranged from 4.7% to 11.6%, the mean difference being 9.6%.
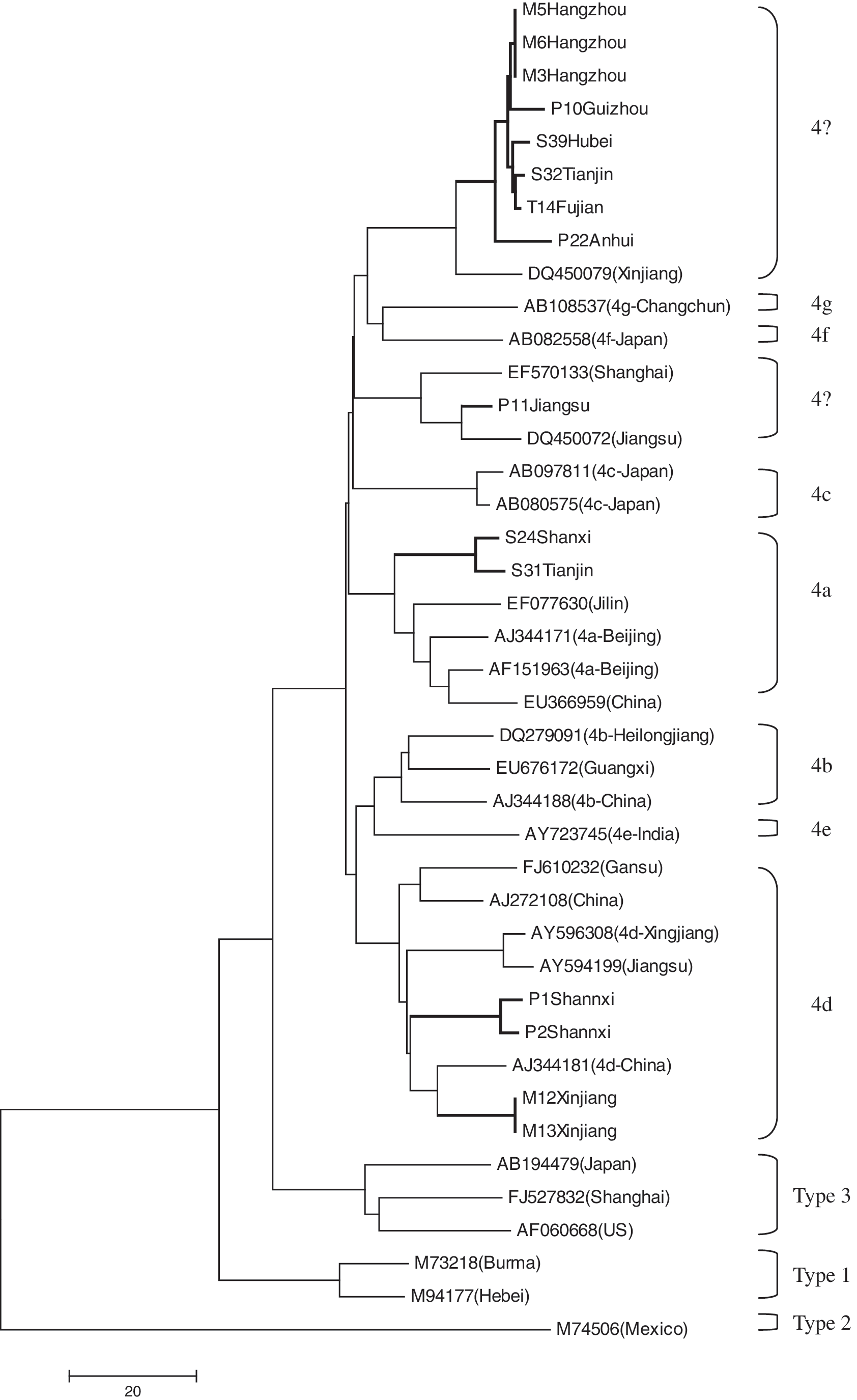
Phylogenetic tree of hepatitis E virus isolates based on partial ORF2 sequences (348 nt). The tree was constructed using the Neighbor-Joining method by Mega 4.1 software. The scale bar indicating the number of character state changes is proportional to the genetic distance. Hepatitis E virus isolates from this study are indicated by thick lines.
Discussion
Hepatitis E is considered by many to be a zoonotic disease, and domestic pigs have been confirmed to be reservoirs of HEV genotypes 3 and 4 (Meng 2009). Our previous investigations showed anti-HEV antibody prevalence in pigs from the northeast of China (including three provinces: Heilongjiang, Jilin, Liaoning) and from Hunan to be 81.6% and 68.3%, respectively (Li et al. 2008, Yu et al. 2009). A similar result was obtained in the present study. The average positivity rate of anti-HEV antibody in swine serum samples from 25 different provinces was 82.2%. The positivity rate varied greatly from province to province, ranging from 10.9% to 100%. Because samples were collected from only a few farms in each province, this variability may reflect differences among farms rather than among provinces. The age of pigs, breeding environment, and other factors can affect the positivity rate of HEV in a farm (Li et al. 2009a, 2009b). Therefore, these results indicate that HEV is prevalent in pig herds across China and the prevalence is relatively high overall.
Mammalian HEV is divided into four genotypes, and these genotypes have a distinct geographic distribution (Okamoto et al. 2007). A candidate fifth genotype was described recently (Zhao et al. 2009). Genotype 4 is the most common in patients with sporadic hepatitis E and in pigs in China, including Hong Kong (Wang et al. 2002, Lam et al. 2009, Tai et al. 2009). Although each HEV genotype tends to diverge to different subtypes, there is no consensus as to how many subtypes each genotype contains. Further, new variants are constantly being discovered, especially within genotypes 3 and 4. On the basis of the HEV sequences currently deposited in the GenBank database, HEV genotype 4 comprises at least seven subtypes, named 4a–4g (Lu et al. 2006). The subtypes 4a, 4b, 4c, 4d, and 4g include Chinese strains isolated from humans or swine. Candidate new subgroups, which are clearly different from these seven subtypes, were found in Hunan province and in northeast China (Li et al. 2008, Yu et al. 2009). To understand completely the genetic diversity and distribution of swine HEV in China, representative strains from each subtype of HEV genotype 4 described in Lu's report, along with the full-length Chinese swine HEV isolates in GenBank, were used to construct a phylogenetic tree using the isolates of this study. Nine subgroups were found within genotype 4, including subgroups 4a–4g identified previously by Lu et al. (2006) and two new subgroups. The 15 swine isolates from the current study were dispersed among 4a, 4d, and the two new subgroups. Strains identified in the same region tend to cluster together. However, one region may harbor more than one subgenotype as described previously (Li et al. 2008, Yu et al. 2009), and each subgroup may contain strains with similar sequences from distinct regions. For instance, in this study, six strains from five different provinces clustered clearly to form a new subgroup, while one Jiangsu isolate clustered with two other previously reported isolates to form another new subgroup. These results indicate that genotype 4 is predominantly prevalent among pigs, and genotype 4 HEV is highly diverse in China. However, the geographic heterogeneity of swine HEV seems becoming blurred. One probable reason is that pig trading across the country leads to the rapid spread of HEV strains from one region to another.
Genotype 3 HEV also has been implicated as a possible zoonotic agent and may be responsible for sporadic human cases. On the basis of the sequences reported previously, genotype 3 probably originated in European pigs, which were later exported around most of the world (Lu et al. 2006). Genotype 3 HEV strains have been isolated in Taiwan and were considered to be imported from the United States through pig trading (Wu et al. 2002). Recent studies report that this genotype is present exclusively in pig farms scattered in the suburbs of Shanghai, China (Ning et al. 2007, 2008). However, no genotype 3 strain was identified in the present study.
HEV antibody was detected in goat and cattle samples collected from various provinces, the prevalence and titers in both being significantly lower than in pigs in this study. The average HEV prevalence in goats and cattle were 10.4% (73/700) and 28.2% (257/912), respectively. However, anti-HEV positivity rates in some populations of goats and cattle were significantly higher than the average. Twelve of 25 (48%) goat serum samples from Shanxi and 13 of 14 (92.8%) cattle serum samples from Guangdong were positive for anti-HEV antibody. Although no HEV RNA was recovered from the goat and cattle using RT-PCR, some serum samples were HEV antigen positive. These results indicate that cattle and goats may also be infected by HEV and are likely to be potential reservoirs of HEV, as are pigs. Because cattle and goats are popular domestic animals of China and most peasants raise them in the backyard, they have more opportunity to come into contact with people than do pigs. Hence, we cannot neglect their potential roles in the transmission of HEV to humans. A possible reason for our inability to amplify HEV nucleic acid from goats and cattle is that the genomic sequences of HEV in these animals are quite likely divergent from those in pigs and humans, and thus the PCR primers based on swine and human HEV genomes do not bind the goat or cattle HEV RNA efficiently. As noted, none of the antigen-positive goat or cattle samples was positive for HEV RNA. Besides the possibility that the animals were infected with divergent genotypes of HEV, the possibility of false positivity in the antigen tests should be addressed. If this is the case, the antigen test should be improved further.
In conclusion, our study showed that HEV infection is common among pigs in China, genotype 4 HEV being the most common and highly diverse. Goats and cattle also may be infected with HEV, albeit the prevalence being significantly lower than in pigs and no viral RNA detected in them. One possible reason for our inability to detect HEV RNA in cattle and goats is that HEV genotypes from different animal species may have divergent sequences.
Footnotes
Acknowledgments
We thank Mr. Zixin Qiu from Wantai Biopharmaceutical Company (Beijing, China) for providing the HEV antibody and antigen ELISA kits. This study was supported by the “863” project (grant 2006AA02Z453) from the Ministry of Science and Technology, China.
Disclosure Statement
The authors have no potential conflicts of interest.