
Abstract
We report the first molecular evidence of the presence of Babesia sp. EU1 and Babesia microti in Ixodes ricinus ticks in Belgium. A 1-year national survey collected 1005 ticks from cats and dogs. A polymerase chain reaction technique amplifying a part of the 18S rRNA gene detected Babesia spp. in 11 out of 841 selected and validated tick extracts. Subsequent sequencing identified Ba. microti (n = 3) and Babesia sp. EU1 (n = 6). This study has demonstrated a low infection rate (1.31% with 95% CI: 0.65–2.33) of Babesia spp. carriage in I. ricinus ticks in Belgium but, for the first time, reports two potentially zoonotic species belonging to this genus. Coinfection with Ba. microti and Borrelia burgdorferi sensu stricto also was demonstrated. In addition, this study clearly demonstrates that inhibitors of polymerase chain reaction amplification are present in engorged ticks.
Introduction
Human babesiosis, a malaria-like disease, has broad spectrum of clinical presentations from a subclinical infection to death according to age, immunocompetence, and comorbidity. Asplenic or immunocompromised patients represent the most susceptible groups. In Europe, human infections acquired from ticks and also from blood transfusions (Krause 2003) are rarely detected but potentially fatal (Homer et al. 2000, Hunfeld et al. 2008).
Man is an accidental host, and before the molecular era, most European human infections were attributed to Babesia divergens, the causative agent of cattle babesiosis in temperate countries (Herwaldt et al. 2003). More recently, a new candidate species designated Babesia sp. EU1 (for Europe 1 and proposed as Ba. venatorum) was described for the first time in two asplenic patients (Herwaldt et al. 2003) and also in roe deer in Europe (Duh et al. 2005, Bonnet et al. 2007). This species seems to be closely related to Babesia odocoilei (a white-tailed deer parasite) and phylogenetically in a sister group with Ba. divergens (Herwaldt et al. 2003). Babesia microti, a parasite of small mammals, is the main agent of human babesiosis in North America. This parasite also occurs in ticks in Europe (Duh et al. 2003, Hartelt et al. 2004, Nijhof et al. 2007). However, only two cases of human babesiosis attributed to Ba. microti have been reported in Europe (Meer-Scherrer et al. 2004, Hildebrandt et al. 2007) and one of these cases remains controversial (Gray 2006). In the United States, other zoonotic Babesia spp. and Babesia-like pathogens have been identified using molecular tools: WA1 (for Washington 1) (Quick et al. 1993) and MO1 (for Missouri 1) (Herwaldt et al. 1996).
In Belgium, there is little information regarding the prevalence of the different species of Babesia and their potential zoonotic impact. Ba. divergens is endemic in the southern part of the country (Famerée et al. 1977, Losson 1989) and was considered absent from other parts of Belgium (Saegerman et al. 2007), although, recently, an outbreak of cattle babesiosis was reported in Vlaams Brabant (Central Belgium). Introduction was probably through importation of infected cattle from the south of Belgium (Everaert et al. 2007). As far as we are aware, human babesiosis due to Ba. divergens has never been reported in Belgium.
In Belgium, Ixodes ricinus is the most prevalent tick species and an important vector of diseases such as babesiosis, borreliosis, and anaplasmosis. It is a competent vector of both Ba. microti and Babesia sp. EU1 (Foppa et al. 2002, Becker et al. 2009, Bonnet et al. 2009). Dogs and cats are close to humans and have often been proposed as effective sentinel animals to assess the risk of human tick-borne diseases (Olson et al. 2000, Duncan et al. 2005).
This study was designed to assess by means of molecular tools the presence of and the infection rate of potentially zoonotic Babesia species in different species of ticks found on dogs and cats in Belgium.
Materials and Methods
From April 2008 to April 2009, a nationwide survey was performed in Belgium. In the 25 veterinary districts, three companion animal practices were selected by stratified randomization. Ticks from cats and dogs, randomly submitted to the 75 veterinary practices, were collected and preserved in 80% alcohol. The ticks were morphologically identified up to stage and species level using a standard key for morphological identification (Arthur 1963). The sex and repletion level were recorded for adults. For each animal enrolled in the study, information was available via a questionnaire completed by the veterinarian about location, date of collection, animal description (dog/cat, age, sex, and breed), and life habits. Ticks collected from dogs and cats with a recent travel history (3 weeks or less before submission) were excluded from the trial. One tick per animal was selected, but when several tick species were present on a given animal, one tick of each species was randomly selected for further analysis.
Tick DNA extraction was performed using a protocol with proteinase K (20 mg/mL) (Boom et al. 1990).
To detect false-negative results due to polymerase chain reaction (PCR) inhibition and to validate the efficiency of the DNA extraction, an additional PCR targeting a 325-bp DNA fragment corresponding to the tick 16S rRNA gene was included. This PCR was set up using 16S + 1 and 16S − 2 primers (Baumgarten et al. 1999). Only positive samples were further analyzed for the presence of Babesia spp.
Repletion level was recorded and potential inhibition due to engorgement of the ticks and the effect of DNA dilution on results was assessed. To do this, 335 DNA samples were chosen randomly and diluted 1/10. Any negative sample was then tested either neat or after 1/100 dilution. Remaining negative samples were again diluted to 1/1000. DNA concentration was recorded.
A Babesia spp. genus-specific PCR was developed according to Casati et al. (2006) using BJ1 and BN2 primers and based on the amplification of a 411–452-bp fragment of the multicopy 18S rRNA gene. To assess the specificity and to validate this PCR method, the following positive controls were used: Babesia canis extracted from canine blood (kindly supplied by ENV), Ba. divergens extracted from cattle blood and Babesia sp. EU1 from deer blood (both supplied by ENV), and Babesia sp. EU1 (supplied by Zurich University). Whenever a tick was found positive for Babesia spp., the other ticks found on the same animal were also examined.
To assess the sensitivity of the Babesia spp. PCR, the DNA concentration of an extract originating from a Ba. canis strain (ENV) was measured by spectrometry (Nanodrop; Thermo Scientific). The extract was then serially diluted 10-fold with sterile water.
Tick DNA samples positive for Babesia spp. were sequenced from PCR products and after cloning. The PCR products were purified using MSB-Spin PCRapace kit (Invitek) and cloned with Topo TA Cloning® Kit for sequencing (Invitrogen). Cycle sequencing reactions were performed by BigDye terminator v3.1 (3730 DNA analyzer; Applied Biosystem) by Giga Genomics Facility (Liège University, Belgium). DNA sequencing was performed in both directions.
A consensus sequence was made according to the results of sequencing of the PCR products and the cloned PCR products via BioEdit program (Hall 1999).
The consensus sequences were analyzed using BLASTn searches in GenBank (
Phylogenetic analysis was performed from consensus and reference sequences by using the neighbor-joining method with Mega 4 program (Tamura et al. 2007). The support for the tree nodes was calculated with 1000 bootstrap replicates.
Ba. microti-positive samples were analyzed for potential coinfection with Borrelia spp. following a PCR protocol amplifying a part of the ospA gene (Demaerschalck et al. 1995). Plasmid containing genomic DNA of Borrelia afzelii (kindly supplied by Utrecht University) was used as positive control. Samples positive for Borrelia spp. were sequenced. Cycle sequencing reactions were performed using the Sanger method by the Medical Centrum Genetica (Ghent University Hospital, Belgium).
Results
Among the ticks collected from 561 dogs and 416 cats, four tick species were present: Ixodes hexagonus, Ixodes ricinus, Dermacentor reticulatus, and Rhipicephalus sanguineus (Table 1).
Eight hundred forty-one out of the 1005 tick extracts were selected and validated by PCR amplifying the 16S rRNA tick gene.
Of the 335 samples tested for inhibitors, a DNA dilution factor of 1/10 most adequately reduced the PCR inhibition. Without dilution, 85% of samples tested were inhibited. With a 1/10 dilution, 80% of the 335 tick extracts were positive. Engorgement had an effect. At a 1/10 dilution, 82% (n = 55) of the negative samples were collected from engorged or semiengorged ticks, whereas only 11% (n = 32) of the positive samples were collected from engorged ticks. In some samples negative at a 1/10 dilution, increasing the dilution factor to 1/100 improved the sensitivity and some tested positive at a 1:1000 dilution (Fig. 1).

Influence of the DNA dilution factor on the 16S rRNA gene tick polymerase chain reaction inhibition. Nt, number of samples tested; Nrt, number of samples tested randomly; Np, number of positive samples; Nn, number of negative samples. *For one sample the test was not performed.
The minimal DNA concentration allowing the detection of the 411–452 bp was estimated at 94 pg/μL (data not shown).
Eleven samples out of the 841 selected and validated DNA extracts were positive for Babesia spp; all were I. ricinus giving an infection rate estimated at 1.31% (95% CI: 0.65–2.33). These infected ticks had been collected from healthy cats and dogs, all living in rural areas and allowed to walk freely in gardens, wooded areas, and meadows. None of the other ticks from these same animals were positive.
Positive samples were located throughout Belgium (Fig. 2).

Map of Belgium showing the localities where Babesia spp. were found. A, Beveren; B, Ravels; C, Beerse; D, Herentals; E, Zichem; F, Bruyelles; G, Spontin; H, Ciney; I, Theux; J, Arbrefontaine. Two Babesia sp. EU1 were found in Theux.
In positive samples, PCR products were sequenced and results are available for seven. The 11 positive samples were cloned: 2 samples could not be cloned, whereas 7 produced results (including 2 samples found negative through direct sequencing). Following the consensus sequences, three samples (accession no. GQ856649, GQ856652, and GQ856653) had 99% homology with Ba. microti (accession no. AF231349) and six others (accession no. GQ856650, GQ856651, GQ856654, GQ856655, GQ856656, and GQ856657) with Babesia sp. EU1 (accession no. AY046575).
A single Ba. microti-positive sample (found in a nymph from Herentals) was also positive for Borrelia burgdorferi sensu stricto.
A phylogenetic tree including each of the nine sequenced Babesia samples and some reference sequences is given in Figure 3.
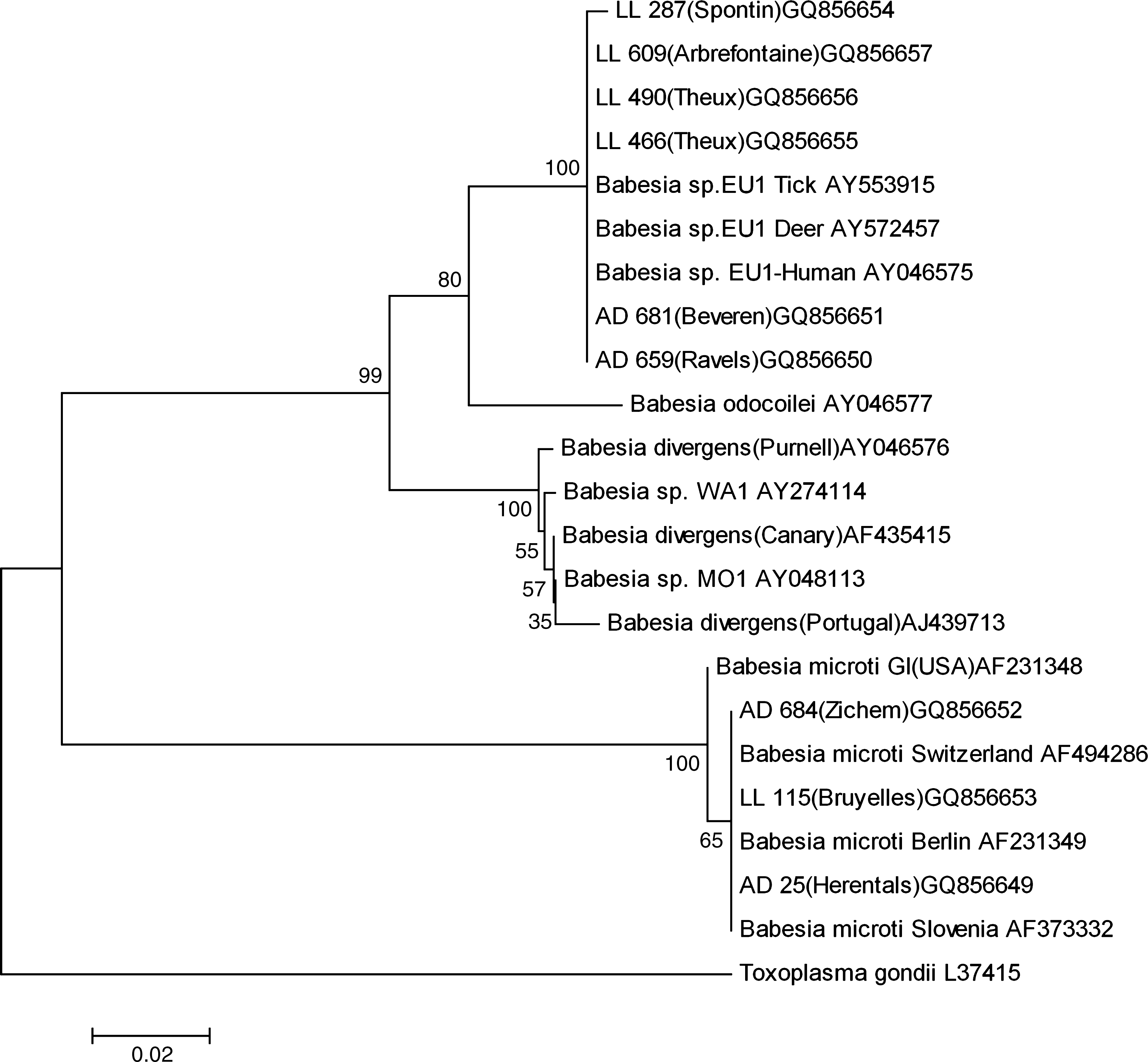
Phylogenetic tree based on a fraction (411–452 bp) of 18S rRNA gene of selected Babesia spp. The accession numbers for the reference sequences follow the corresponding names in the tree. Samples are identified with a code (2 letters and number); geographical localizations are given in brackets followed by the GenBank accession number.
Discussion
This study, based on molecular identification, shows an infection rate with Babesia spp. in I. ricinus in Belgium of 1.31% (95% CI: 0.65%–2.33%), which is in agreement with the data from nearby countries, that is, the Netherlands (0.6%–2.3%) (Nijhof et al. 2007, Wielinga et al. 2008), southern Germany (1%) (Hartelt et al. 2004), and Switzerland (0.7%–1.7%) (Casati et al. 2006). In central and eastern European countries, infection rates reach much higher values: 16.3% in Poland (Skotarczak and Cichocka 2001), 9.6% in Slovenia (Duh et al. 2001), and up to 51% in Austria (Blaschitz et al. 2008). These marked differences could be due to environmental factors controlling tick density. Also, the selected collection technique (from an animal host or from the environment through flagging) could also influence detection rate.
This study, based on the amplification of a fragment of the 18S rRNA gene, has demonstrated, for the first time, Ba. microti and Babesia sp. EU1 in I. ricinus ticks collected from pets throughout Belgium. Both species were known to occur in several European countries (Casati et al. 2006, Bonnet et al. 2007, Wielinga et al. 2008). Taking into account the fact that other ticks collected from a pet carrying a positive tick were all negative, no conclusion can be drawn about transmission through cofeeding.
The following questions remain: Is Babesia sp. EU1 A new species? What are exactly the etiologic agents of human babesiosis in Europe?
Based on morphological criteria, serological cross-reactivity (Herwaldt et al. 2003), vector identity (Becker et al. 2009, Bonnet et al. 2009), and host range, Ba. divergens and Babesia sp. EU1 could belong to the same species.
Until now, the main differences between Ba. divergens and Babesia sp. EU1 are based on a 31 bp difference on the 18S rRNA gene and the inability of Babesia sp. EU1 to infect and kill gerbils (Meriones unguiculatus) after experimental infection (Herwaldt et al. 2003). The phylogenetic tree (Fig. 3) is in agreement with other authors' finding: Babesia sp. EU1 is close to Ba. odocoilei and is in a cluster group with Ba. divergens (Häselbarth et al. 2007). Unfortunately, in Europe, very few molecular data are available from confirmed human cases of babesiosis (Olmeda et al. 1997), and consequently, in many cases, the accurate identity of the responsible pathogen is unknown.
It remains unclear why the epidemiology of Ba. microti seems different in the United States (where zoonotic impact is well known) and Europe (where confirmed human cases are very rare). Some hypotheses exist: these differences might be due to the circulation of different strains as suggested by the phylogenetic tree or due to the fact that others tick species are the vectors in the United States and Europe; and possibly, in Europe, the real prevalence in humans is underestimated because of subclinical infections (Persing et al. 1992) or coinfection with other tick-borne pathogens such as Borrelia spp. (Krause et al. 2002). This coinfection has again been demonstrated in the present animal survey. Further, the lack of specific diagnostic tools may explain the paucity of information about this group of pathogens in Europe.
In this study, ticks collected from dogs and cats were examined. These two species live in close contact to humans and are exposed together with their owners, through travel and leisure activities, to I. ricinus ticks and the pathogens they carry. In this respect, pets could be considered as sentinels for tick-borne diseases such as borreliosis, anaplasmosis (Olson et al. 2000, Duncan et al. 2005), and babesiosis. This seems to be confirmed in this survey at least for the genus Babesia.
Several compounds in blood have been suggested to be PCR inhibitors, such as hemoglobin, lactoferrin, and immunoglobulin G. Hemoglobin and lactoferrin contain iron; the inhibitory effects of both proteins may be related, in part, to their ability to release iron ions (Al Soud and Radstrom 2001). When they feed, ticks ingest all of these components. Concerning our investigations, inhibition occurs in most cases when ticks were engorged. We assumed that these PCR inhibitions were probably due to these blood compounds.
Footnotes
Acknowledgments
The authors thank Prof. Gilles Bourdoiseau (ENV Lyon), Dr. Sarah Bonnet (INRA Nantes), Dr. Alexander Mathis (University of Zurich), and Prof. Frans Jongejan (Utrecht University) for kindly supplying the positive controls. The authors are very grateful to Axel Mauroy and Catherine Généreux for their scientific support and Françoise Maréchal for her technical support. The authors thank the veterinary practices for their participation in this survey and acknowledge the financial support of Bayer Animal Health Belgium.
Disclosure Statement
No competing financial interests exist.