
Abstract
Thirty-two strains of Borrelia burgdorferi sensu lato were isolated from Ixodes persulcatus ticks collected from northeastern China from May to June in 2004 and 2005. Restriction fragment length polymorphism (RFLP) analysis and sequence analysis of 5S-23S rRNA intergenic spacer revealed that 29 (90.6%) belonged to Borrelia garinii, demonstrating B, C, and a unique pattern. The remaining three isolates (9.4%) were Borrelia afzelii with pattern D. The phylogenetic analysis based on 5S-23S rRNA intergenic spacer showed that B. garinii and B. afzelii genospecies clustered into two separate lineages. B. garinii strains were classified into three different branches: All the strains with RFLP pattern C were in the same branch, strain VH10 with a unique RFLP pattern clustered with strains VH9 and MDH2 with pattern B, and the rest of the strains with pattern B constitute another branch. These findings demonstrate the genetic diversity of B. burgdorferi sensu lato isolates from northeastern China.
Introduction
Lyme borreliosis in mainland China was first reported in 1986 in Heilongjiang province, which is located in northeastern China (Ai et al. 1988). Thereafter, the extensive epidemiological surveys carried out from 1987 to 1996 reported the existence of Lyme borreliosis in at least 17 provinces throughout the country (Zhang et al. 1997). Among them, the forest regions of northeastern China were considered to be seriously endemic for Lyme borreliosis (Zhang et al. 1997). Ixodes persulcatus was proved to act as the major vector in this area (Wan et al. 1998). Two species, B. garinii and B. afzelii, were discovered in this area by restriction fragment length polymorphism (RFLP) analysis of 16S-23S rRNA gene and 5S-23S rRNA intergenic spacer (Liang and Zhang 1996, Zhang et al. 1997, Takada et al. 1998). Whether B. burgdorferi sensu stricto exists in northeastern China has been disputed (Liang and Zhang 1996, Zhang et al. 1997, Takada et al. 1998). Since 1998, few epidemiological surveys in this area have been reported. However, many studies recently performed in other areas of mainland China provided evidences of new B. burgdorferi sensu lato species such as Borrelia sinica sp. nov. and Borrelia yangtze sp. nov. by using more fine-scaled molecular techniques (Masuzawa et al. 2001, 2004, Chu et al. 2008). We, thus, hypothesize that northeastern China, as one of the highly endemic regions for B. burgdorferi, might possess more diversity of B. burgdorferi sensu lato population than that previously reported (Liang and Zhang 1996, Zhang et al. 1997, Takada et al. 1998). From 2004 to 2005, a survey for B. burgdorferi sensu lato in ticks was carried out in northeastern China; the genetic diversity of B. burgdorferi sensu lato population was assessed by jointly using RFLP analysis and phylogenetic analysis of 5S-23S rRNA intergenic spacer.
Materials and Methods
Sample collection and Borrelia isolation
A total of seven survey sites were chosen in the forested highlands in the Inner Mongolia Autonomous Region, Heilongjiang Province, and Jilin Province in northeastern China, which are documented to be the endemic foci of Lyme borreliosis (Fig. 1). Two representative landscapes (taiga coniferous forest and mixed forest of larch and birch) were sampled in the region. From May to June in 2004 and 2005, ticks were collected by flagging over vegetation. After identifying the species and developmental stage, they were kept alive in moist bottles until use (Teng 1991).
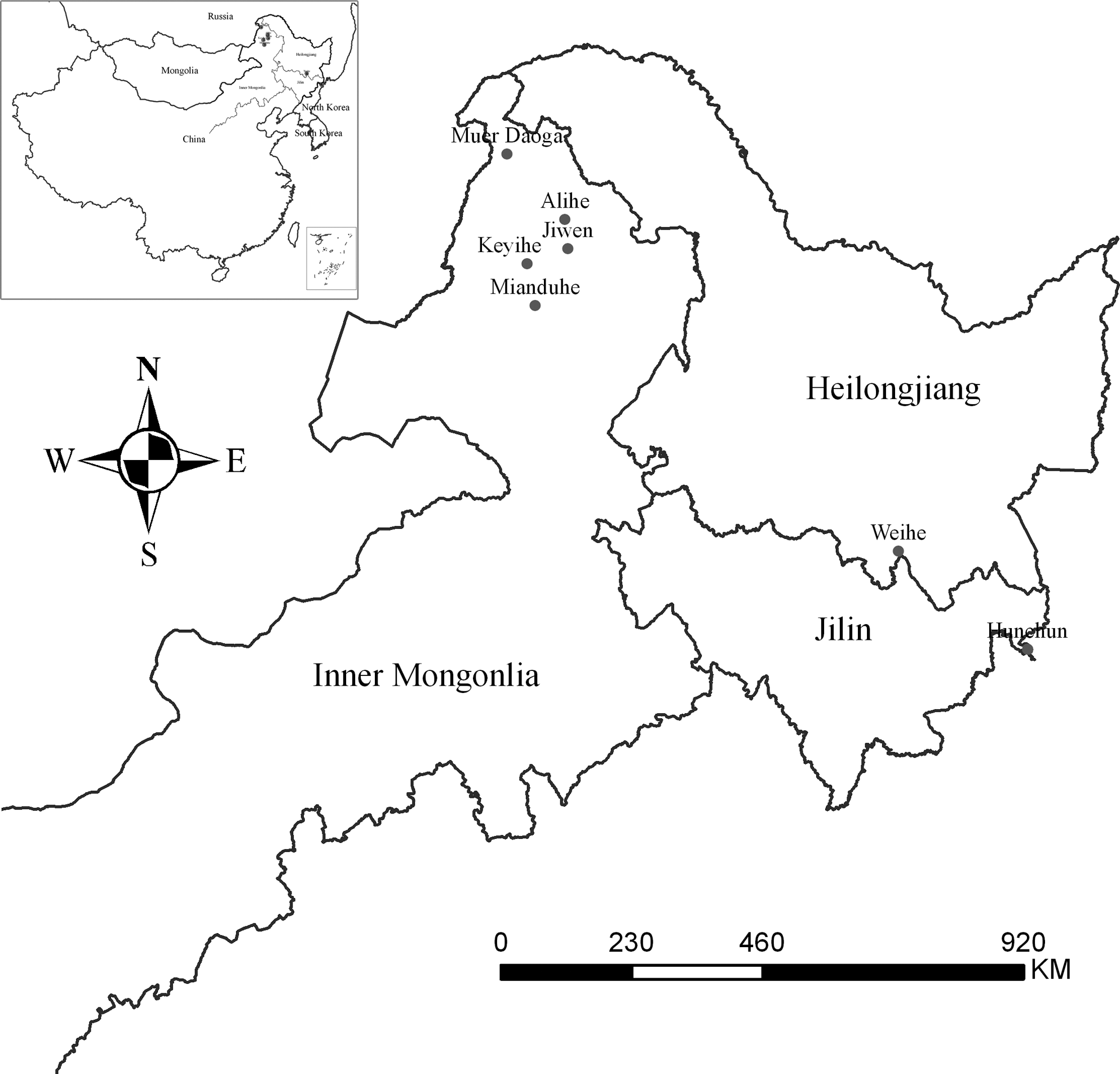
Survey sites in northeastern China.
Unfed adult ticks were used for B. burgdorferi sensu lato isolation. Twenty ticks of the same species were pooled, disinfected in 75% ethanol for 20 min, and washed thrice in sterile phosphate-buffered saline. Then, the ticks were dried on sterile filter paper, ground, and inoculated into 2 mL of Barbour-Stoenner-Kelly H medium (Sigma, St. Louis, MO) supplemented with 6% rabbit serum and 1% antibiotic mixture for Borrelia (Sigma). The cultures were kept in tightly capped tubes, incubated at 29°C, and examined once a week by dark-field microscopy.
DNA extraction
DNA extraction from Borrelia strains was performed as previously described (Postic et al. 2007). Briefly, Borrelia cultures were harvested by centrifugation (10,000 g; 20 min). The bacterial pellet was washed in phosphate-buffered saline and resuspended. The DNA was extracted from the centrifugation pellet of cultivated isolates by boiling in water at 100°C for 10 min and stored at −20°C until use.
Polymerase chain reaction
A primer set (primer 1 [5′-CTGCGAGTTCGCGGGAGA-3′] and primer 2 [5′-TCCTAGGCATTCACCATA-3′]), previously described by Postic et al. (1994), was applied to amplify the variable spacer between two conserved sequences, the 3′ end of the 5S rRNA and the 5′ end of the 23S rRNA, which was expected to yield 226 to 266 bp fragment. The polymerase chain reaction (PCR) amplification was performed in a volume of 20 μL containing 15.8 μL deionized water, 1 μL DNA template, 2 μL 10 × PCR buffer (containing 100 mM Tris-HCl, 500 mM KCl, and 15 mM MgCl2), 0.2 μL Taq DNA polymerase (5 U/μL), 0.2 μL deoxynucleoside triphosphate mix (10 mM) (all from TaKaRa, Shuzo Co. Ltd., Kyoto, Japan), and 0.4 μL of each primer (12.5 μM). The cycling conditions for the amplification involved a 2-min denaturation at 96°C, followed by 30 cycles of 94°C for 30 s, 59°C for 30 s, and 72°C for 40 s, and a final extension at 72°C for 5 min. In parallel with each amplification, a negative control (distilled water) was included.
All the PCR products were separated by 1.5% agarose gel electrophoresis, stained with ethidium bromide, and visualized under UV light. To minimize contamination, DNA extraction, the reagent setup, amplification, and agarose gel electrophoresis were performed in separate rooms.
RFLP analysis
The amplification products of 5S-23S rRNA intergenic spacer were further analyzed by RFLP. Previously extracted DNA of B. garinii, B. afzelii, and B. burgdorferi sensu stricto were used as positive controls. For each strain, 13 μL amplified DNA was digested at 37°C overnight with endonucleases MseI (New England Biolabs, Beverly, MA) according to the manufacturer's recommendations. Electrophoresis was conducted in 16.0% polyacrylamide gel at 100 V for 3 h. The gels were stained with silver staining, and bands were subsequently visualized under white light. Fifty base pair DNA Ladder Marker (TaKaRa, Shuzo Co. Ltd.) was used as a molecular mass marker. Genospecies of B. burgdorferi sensu lato was identified according to RFLP profiles of each sample.
DNA sequencing and sequence analysis
The 5S-23S rRNA intergenic spacer amplicons were gel purified by using Qiaquick Gel Extraction Kit (Qiagen, Hilden, Germany) and sequenced by a dideoxynucleotide cycle sequencing method with an automated DNA sequencer (ABI Prism 377; Perkin-Elmer, Foster City, CA). The sequences determined in this study have been deposited to GenBank database under the accession numbers listed in Figure 3. The exact length of restriction fragments was obtained through DNA sequence analysis using Vector NTI 9.0 software (Lu and Moriyama 2004). The sequences with unique RFLP patterns were compared with those previously deposited in GenBank using BLAST program from the National Center for Biotechnology Information Web site.
Phylogenetic analysis
The Clustal X software (version 1.8) (Thompson et al. 1997) and PHYLIP software package (version 3.63) (Retief 2000) were used for sequence alignment and phylogenetic analyses, respectively. The distance matrices for the aligned sequences were calculated by the Kimura 2-parameter method. The phylogenetic trees were constructed using the neighbor-joining method (Saitou and Nei 1987). The stability of the tree was evaluated by bootstrap analysis with 1000 replications.
Results
Borrelia isolation
A total of 118 pools of 2360 unfed adult ticks comprising three species, I. persulcatus (75 pools), Dermacentor silvarum (33 pools), and Haemaphysalis concinna (10 pools), were collected and used for B. burgdorferi sensu lato isolation. Thirty-two isolates were obtained from 75 pools of I. persulcatus ticks 4 weeks after inoculation in Barbour-Stoenner-Kelly H cultures (Table 1). Among them, 16 B. burgdorferi sensu lato isolates were obtained from 40 pools of ticks collected in Inner Mongolia Autonomous Region; 13 were from 20 pools collected in Heilongjiang Province and the remaining 3 were from 15 pools of ticks collected in Jilin Province (Table 1). Considering the possibility of multiple infection of the same genotype in one pool of ticks, we could only estimate the minimal prevalence of B. burgdorferi sensu lato in I. persulcatus as 2.13% (32/[75 × 20]). No Borrelia isolate was obtained in either H. concinna from Inner Mongolia Autonomous Region or D. silvarum from Jilin and Heilongjiang provinces.
Borrelia identification
PCR amplifications targeting the 5S-23S rRNA intergenic spacer of B. burgdorferi sensu lato generated fragments ranging from 246 to 255 bp in length in this study. After cleavage by MseI, the PCR products from 32 isolates exhibited four distinct patterns. Of these, 16 (50.0%) isolates generated pattern B of B. garinii, 12 (37.5%) with pattern C of B. garinii, 3 (9.4%) with pattern D of B. afzelii, and 1 (3.1%) with a unique pattern U (Fig. 2 and Table 2). Exact fragment sizes generated after digestion with the restriction enzymes MseI were determined by sequence analysis of the amplicons (Table 2).
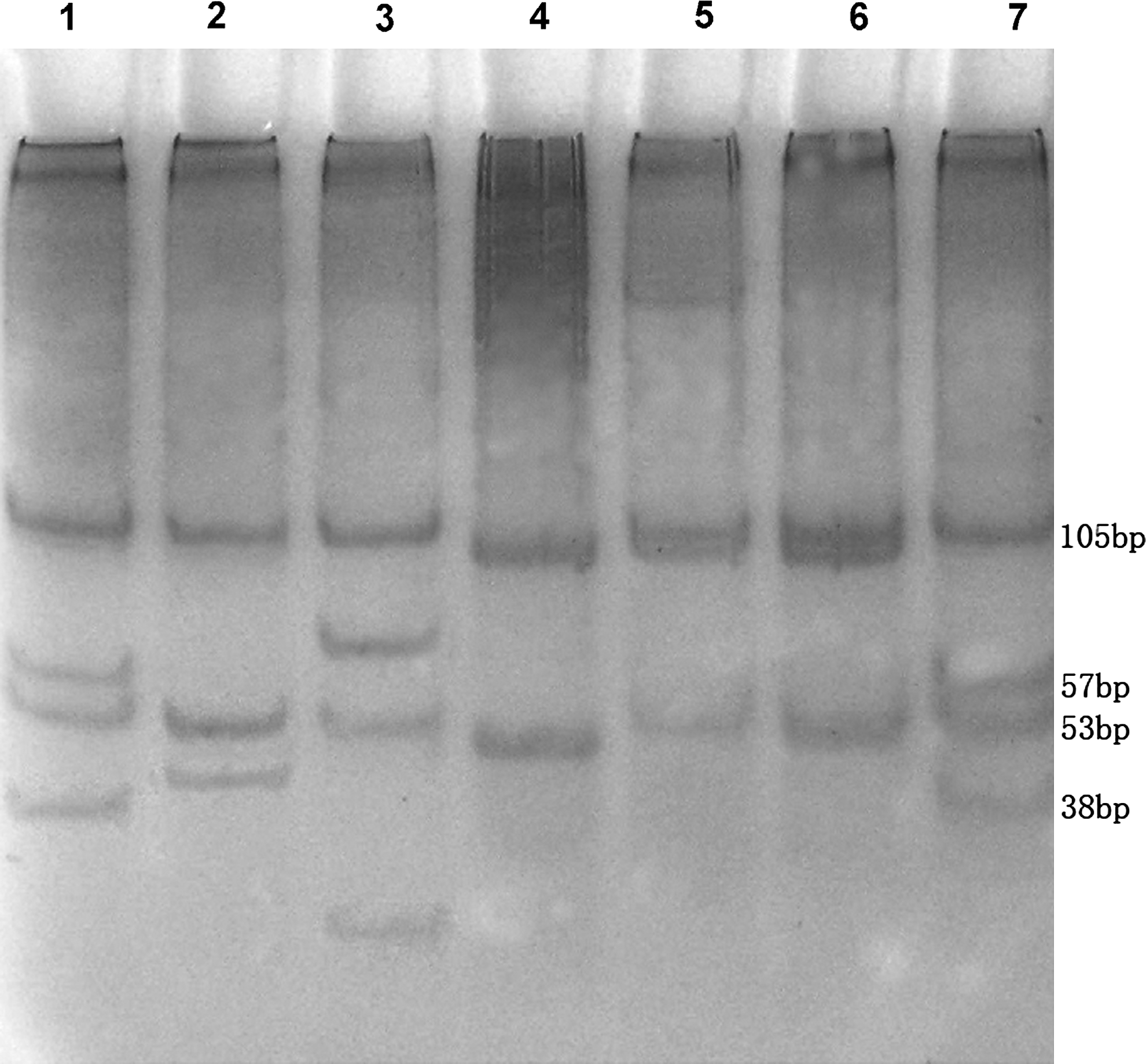
Representative MseI restriction patterns of the 5S-23S rRNA intergenic spacer of Borrelia burgdorferi sensu lato isolates from northeastern China. The molecular size standards are indicated on the right of the gel. Lane 1, pattern C of Borrelia garinii strain JW1; lane 2, pattern U of B. garinii strain VH10; lane 3, pattern D of Borrelia afzelii strain VH7; lane 4, 50 bp DNA marker; lane 5, patterns B of B. garinii strain JW2; lane 6, pattern B of B. garinii strain VH4; lane 7, pattern C of B. garinii strain NMK7.
RFLP, restriction fragment length polymorphism.
The sequence comparison revealed that the 5S-23S rRNA intergenic spacer of the isolate VH10 with a unique RFLP pattern displayed the highest similarity (99.6%) with that of B. garinii strain Ip-1741 (GenBank accession no. AM748062) isolated from Russia. Moreover, the 5S-23S rRNA intergenic spacer sequence of the isolate VH10 was 98.8% similar with those of B. garinii strains Nr168 (GenBank accession no. AB178380) and Ipl-4845 (GenBank accession no. AM748060), both of which were also isolated from Russia and exhibiting pattern B. Sequence analysis indicated that VH10 had a base A at position 58, instead of C as shown in the strain Nr168. This change generated a recognition locus by enzyme MseI (TTAA, position 95–98). Therefore, four fragments (105, 53, 51, and 44 bp) were generated by digestion with MseI, rather than three fragments (105, 95, and 53 bp) for pattern B. Based on the combined analysis of RFLP patterns and sequences, isolate VH10 was classified into B. garinii genospecies. Thus, the 32 isolates could be classified into two genospecies, with 29 (90.6%) belonging to B. garinii and 3 (9.4%) belonging to B. afzelii.
Phylogenetic analysis of 5S-23S rRNA intergenic spacer
The sequence alignment indicated that all B. afzelii strains had the identical sequence of 5S-23S rRNA intergenic spacer, whereas B. garinii strains showed 93.1%–100% sequence similarity within the group. In addition, six groups of strains that showed identical within-group sequence of 5S-23S rRNA intergenic spacer were determined (i.e., isolates HQ and TP; isolates MDH1 and JL2; isolates NMK3, VH1, and ALH1; isolates NMK6, VH3, and VH18; isolates NMS2, VH11, and VH20; isolates VH4 and VH19). Therefore, one representative sequence of each strain group was selected to create the phylogenetic trees. In addition, the 5S-23S rRNA intergenic spacer sequences of standard strains and (or) representative strains of B. burgdorferi sensu stricto, B. afzelii, Borrelia Valsisiana, and Borrelia lusitaniae, and B. garinii strains with different RFLP patterns observed till now, were derived from GenBank and used for comparison in this study.
On the phylogenetic tree, the strains clustered into different branches depending on the genospecies of B. burgdorferi sensu lato, except for B. garinii strains ChY3 and 161 with pattern R, which did not fall into the branch of B. garinii strains but a freestanding branch. The isolates in this study clustered into two separate lineages: B. garinii strains fell into the large cluster with B. garinii strains NT29 and 20047; B. afzelii strains clustered with B. afzelii strains VS461 and 17Y, moderately supported by bootstrap analysis. Further, the cluster of B. garinii strains were in three branches: All of the B. garinii strains with pattern C clustered with Japanese B. garinii strain NT29, B. garinii strain VH10 with a unique RFLP pattern clustered with two B. garinii strains VH9 and MDH2 with pattern B, and the rest of the B. garinii strains with pattern B clustered with European B. garinii strain 20047 (Fig. 3).
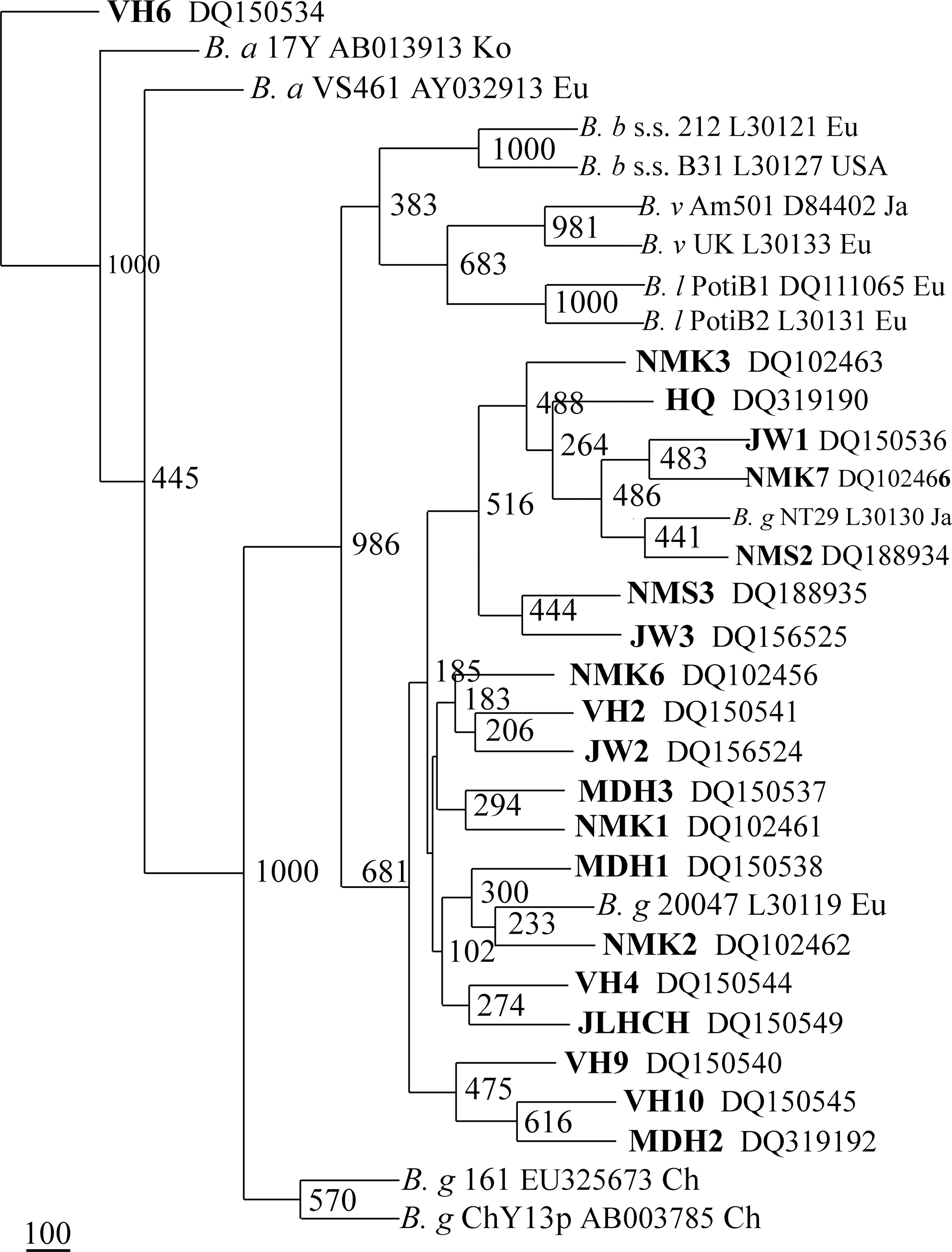
Unrooted neighbor-joining phylogenetic trees based on the 5S-23S rRNA intergenic spacer sequence created by using neighbor-joining method with Kimura two-parameter analysis and bootstrap analysis of 1000 replicates. Isolates in this study are indicated in bold type. GenBank accession numbers and geographic sources of reference strains are marked after each name of strains. Eu, Europe; USA, United States; Ja, Japan; Ko, Korea; Ch, China; B. b s.s., B. burgdorferi sensu stricto; B. b, B. garinii; B. a, B. afzelii; B. v, B. valaisiana; B. l, Borrelia lusitaniae.
Discussion
In this survey, taiga coniferous forest and mixed forest of larch and birch were chosen as sampling sites that ticks prefer to inhabit, to obtain higher abundance and more genetic diversity of B. burgdorferi sensu lato population. A total of three species of ticks, I. persulcatus, D. silvarum, and H. concinna, were collected, and I. persulcatus was the most commonly collected on vegetation. Moreover, B. burgdorferi sensu lato isolates were only obtained from I. persulcatus ticks. These findings are consistent with the previous report of I. persulcatus serving as the principal vector of B. burgdorferi sensu lato in northeastern China (Wan et al. 1998).
In this study, the prevalence (2.13%) of B. burgdorferi sensu lato in I. persulcatus ticks is incredibly low compared with that of 21.93% (293/1336) determined by the PCR test of single ticks in our other epidemiologic study conducted during 2004 and 2005 (Chu et al. 2006). The pool of as many as 20 ticks was used for isolation, instead of the single tick isolation. This was performed mainly to increase the isolation rate of B. burgdorferi sensu lato. However, inevitably, there were more than one infected tick in one pool, and the prevalence might be underestimated. Although our results actually disclosed no mixed culture by PCR-RFLP analysis and sequence analysis of 5S-23S rRNA intergenic spacer, the following two possibilities should be considered: more than one infected tick with the same B. burgdorferi genotype and the outgrowing of one strain over the other in one pool of samples.
Two genospecies, B. garinii and B. afzelii, were found in northeastern China, with B. garinii as the predominant one. No other genospecies was obtained at any survey sites, including Jilin Province where B. burgdorferi sensu stricto was documented to be isolated from I. persulcatus ticks in 1996 (Liang and Zhang 1996, Zhang et al. 1997). B. garinii was isolated in all survey sites, but B. afzelii was isolated only in Weihe county of Heilongjiang Province, where the first case in mainland China was reported. The higher prevalence of B. garinii might be due to the wider range of its suitable ecologic niche (e.g., reservoirs) when compared with B. afzelii (Kurtenbach et al. 2002). On the other hand, it showed that there was the greater genetic diversity of B. burgdorferi sensu lato population in Weihe county of Heilongjiang Province than that in other survey sites. The variation of genetic diversity of B. burgdorferi sensu lato isolates among the different sites might have evolved from ecologic reasons, which deserve further investigation.
Three B. afzelii isolates showed the same patterns by RFLP analysis of 5S-23S rRNA intergenic spacer and also had the same sequence of 5S-23S rRNA intergenic spacer. However, 29 B. garinii isolates exhibited three RFLP patterns, including pattern B, pattern C, and a unique RFLP pattern U, which was not previously reported. Moreover, our other epidemiologic study, which was conducted in Inner Mongolia Autonomous Region of northeastern China during 2004 and 2005, found the presence of B. garinii with another unique pattern R (105, 79, and 53 bp), besides B. garinii with patterns B and C and B. afzelii with pattern D (Chu et al. 2006). However, B. garinii with patterns B, C, and R, as well as B. afzelii with pattern D, was only detected in the previous study in Inner Mongolia in 1996, with the absence of B. garinii with pattern U (Li et al. 1998). These findings displayed the more genetic diversity of B. burgdorferi sensu lato population in northeastern China.
The genetic diversity of B. burgdorferi sensu lato isolates in northeastern China was confirmed by the phylogenetic analysis of 5S-23S rRNA intergenic spacer. Thirty-two isolates clustered into two separate lineages depending on the genospecies in the phylogenetic tree. Moreover, the cluster of B. garinii strains further separated into three different branches. Selection and migration, which are largely host-driven to influence the ecology of Lyme borreliosis, are considered major forces shaping the population structures of B. burgdorferi sensu lato (Kurtenbach et al. 2002).
Notably, B. garinii control strains with unique pattern R (isolate ChY13p and sample 161) fell into a separate cluster, which was close to the cluster of B. afzelii strains but relatively distant from that of other B. garinii strains. However, isolate ChY13p grouped with the other B. garinii isolates with patterns B and C on the phylogenetic tree based on the housekeeping 16S rRNA gene in another study (Li et al. 1998). The different topologic structure of trees might be due to the sequences of different genes chosen: One was the highly variable 5S-23S rRNA intergenic spacer, and the other was the housekeeping 16S rRNA gene. Since the 5S-23S rRNA intergenic spacer is the noncoding region of the pathogen and is not involved in infection or resulting in disease, the phenotypic and genotypic characteristics and the pathogenicity to humans of these unique B. garinii strains need to be further studied, to determine whether the genetic diversity of B. burgdorferi sensu lato population should be considered when designing vaccines and treating patients.
Footnotes
Acknowledgments
The authors are grateful to Dr. Yi Sun for identification of ticks and to Shan-Hu Chen, Guo-Song Ren, Xiu-Xian Zhang, Jian-Hua Zhu, and Feng-Ying Liu for assistance in field work. The authors also thank two anonymous reviewers of this manuscript for valuable comments and suggestions and Dr. Xiaofeng Frank Yang (University of Indiana) for helpful comments on this manuscript. This study was supported by the National Science Fund for Distinguished Young Scholars (No. 30725032), grants from Natural Science Foundation of China (30600506 and 81001266), and National 973 Project (2010 CB530201).
Disclosure Statement
No competing financial interests exist.