
Abstract
Raccoons (Procyon lotor) live at high densities, often in close association with people, in urban areas in Ontario and have been implicated as potential reservoirs of numerous zoonotic disease agents. We collected 137 blood samples from 61 apparently healthy raccoons in a small area of Toronto, Ontario, from June to October 2007 as part of a longitudinal study to determine the seasonal patterns of seroprevalence of Francisella tularensis, avian influenza, and Leptospira. In addition, we collected 35 urine samples by cystocentesis from 23 animals to look for evidence of urinary shedding of Leptospira. All samples were serologically negative for F. tularensis and avian influenza. Nineteen of 61 animals (31%) were positive for Leptospira antibodies in one or more trapping periods. The seroprevalence of Leptospira increased from 5% in June to 38% in October. Of the 19 positive animals, 14 were seropositive for serogroup Grippotyphosa, 4 for serogroup Pomona, and 1 for both serogroups Australis and Grippotyphosa. Raccoons were seronegative to serovars representative of serogroups Autumnalis, Canicola, Icterohaemorrhagiae, and Sejroe. Only one urine sample was culture positive for Leptospira (2.9%). Although we found evidence that raccoons in this study were exposed to leptospires belonging to serogroup Grippotyphosa, likely serovar Grippotyphosa, during the summer and able to shed leptospires in urine, further work is required to determine the importance of raccoons as reservoirs of Leptospira in Ontario.
Introduction
In Ontario, raccoons have been implicated as reservoirs of Leptospira serovar Grippotyphosa for humans and dogs based primarily on serological evidence of exposure to Leptospira (Warshawsky et al. 2000, Prescott 2008). However, Haydon et al. (2002) defines a reservoir as “one or more epidemiologically connected populations or environments in which the pathogen can be permanently maintained and from which infection is transmitted to the defined target population.” Thus, to demonstrate that raccoons are a reservoir of Leptospira, evidence of persistent infection and pathogen shedding, not just pathogen exposure, is required.
The first objective of this study was to determine the seasonal seroprevalence patterns of Leptospira in wild raccoons in an urban area (on the grounds of the Toronto Zoo) and to conduct a longitudinal study to investigate the role of raccoons as reservoirs of Leptospira infections. In addition, we looked for evidence that raccoons on the grounds of the Toronto Zoo were exposed to two other important pathogens: avian influenza A, which has recently been found in raccoons in the United States (Hall et al. 2008), and Francisella tularensis, which has been detected of raccoons in a variety of sites (Berrada et al. 2006).
Materials and Methods
Procedures for trapping and handling raccoons were approved by the Animal Care Committee at the University of Guelph following the guidelines of the Canadian Committee on Animal Care. Forty Tomahawk live traps (Tomahawk Live Trap) were set in two areas on the grounds of the Toronto Zoo (43°49′N, 79°11′W) for three nights each month from June to October 2007. Traps were baited with cat food and set in the evening and checked the following morning. Captured raccoons were brought to a centralized holding area for processing, unless they had already been caught that month, in which case they were released immediately at the point of capture.
Raccoons were anesthetized using an intramuscular injection of 0.05 mg/kg medetomidine hydrochloride (Domitor 1 mg/mL; Pfizer Animal Health, Pfizer Canada) and 5 mg/kg ketamine hydrochloride (Ketaset 100 mg/mL; Wyeth Animal Health) before removal from traps. A numbered metal ear tag (no. 1005-3; National Band and Tag) was placed in one ear and a PIT tag (AVID Canada) was injected subcutaneously between the shoulder blades for subsequent identification. Sex, age class (adult or juvenile, based on animal size and teeth wear/staining), and mass were recorded for each animal. Blood (∼5 mL) was collected from the jugular vein. Blood samples were kept cool during transport back to the laboratory, and serum was extracted and stored frozen at −80°C before serological testing. For leptospiral culture, we attempted to collect urine samples via cystocentesis if the bladder was palpable or from trays placed under the traps during processing. For transport back to the laboratory, 0.5 mL of urine was added to 4.5 mL of sterile 1% bovine serum albumin (BSA) (Ellinghausen 1973) containing 100 μL of 5-fluoruracil/mL. The samples were kept cool during transport to the laboratory and processed on the same day of collection or held overnight at 4°C. After sampling was completed, animals were given an anesthetic reversal agent, 0.25 mg/kg atipamazole (Antisedan 5 mg/mL; Pfizer Animal Health, Pfizer Canada) and placed back in the traps to recover from the anesthetic before release at point of capture.
As part of separate on-going management procedures at the Toronto Zoo, the majority of raccoons were sterilized (vasectomy, or tubal ligation), vaccinated for distemper and rabies, and treated for parasites upon first capture. Nine animals were given penicillin G (1 mL per animal, Benzathine and Procaine Sterile Aqueous Suspension; Vetoquinol Canada) as a precaution against infection after surgery in June. Animals that underwent surgery were released at point of capture the following morning.
A microaggulutination (MA) test as described by Sato et al. (1990) was used to screen serum samples from raccoons for antibodies to F. tularensis. The MA test was a miniaturized form of the tube agglutination test performed in a 96-well-plate format (Brown et al. 1980). Briefly, 25 μL of MA diluent (1% normal rabbit serum and 0.4% phosphate-buffered saline) was applied to each well of a U-bottom microtiter plate. Twenty-five microliters of each test serum was mixed with an equal volume of F. tularensis antigen (SCHU strain grown in-house at the National Microbiology Laboratory (NML)) and serially diluted from 1:4 to 1:8192. Plates were gently tapped to mix contents of the wells and then incubated in a sealed container overnight at room temperature. Tests were read the following morning, and agglutination was observed as a diffuse lattice while nonreactive wells appeared as a button of cells. Wells containing the last dilution in which a discernable button could not be seen were considered the end-point titer. As samples were not paired, only serum samples with an end-point titer of 128 were considered positive for F. tularensis (Ohara et al. 1974).
The presence of antibodies against influenza A viruses was examined by agar gel immunodiffusion as described previously (Thayer et al. 2008). Inactivated A/turkey/Wisconsin/66 (H9N2) antigen concentrate, and positive and negative chicken serum were purchased from Charles River Laboratory.
The presence of antibodies against serovar Autumnalis (strain Akiyami A, representing serogroup Autumnalis), serovar Bratislava (strain Jez Bratislava, serogroup Australis), serovar Canicola (strain Hond Utrecht IV, serogroup Canicola), serovar Grippotyphosa (strain Moskva V, serogroup Grippotyphosa), serovar Hardjo (strain Hardjoprajitno, serogroup Sejroe), serovar Icterohaemorrhagiae (strain M20, serogroup Icterohaemorrhagiae), and serovar Pomona (strain Pomona, serogroup Pomona) was examined by microscopic agglutination test (MAT). Reference cultures and positive serum controls were obtained from KIT Biomedical Research (WHO/FAO/OIE Collaborating Centre for Reference and Research on Leptospirosis, Amsterdam). MAT was carried out as recommended by World Health Organization and International Leptospirosis Society (WHO 2003). Samples were screened at 1:50 dilution and end point titers were subsequently determined for reactive samples. The end point is defined as the highest dilution of serum that showed at least 50% agglutination. The titer is the reciprocal of the highest dilution of the sample that reacted with a reference antigen. Samples with titers of 100 or higher were considered positive for the presence of leptospiral antibodies.
Urine samples were diluted 10-fold in 1% BSA (10−1, 10−2) (Ellinghausen 1973). Four tubes of EMJH medium (Johnson and Harris 1967) containing 0.15% agar, 1% BSA, 1% rabbit serum, and 100 μL of 5-fluoruracil/mL were inoculated with 100 and 500 μL aliquots of each dilution and incubated at 29°C (Bolin et al. 1989). The cultures were examined for leptospires by dark-field microscopy at 1, 2, 4, and 6 weeks.
Results
One hundred thirty-seven serum samples were collected from 61 apparently healthy animals trapped between June and October 2007. Thirty three animals were trapped on more than one occasion and six animals were trapped in all five trapping sessions (Table 1).
All samples were serologically negative for F. tularensis and avian influenza. We had sufficient samples (61) to be 95% confident of detecting antibodies to F. tularensis and avian influenza at our site if the seroprevalence of either agent was >4% (sample size for detection of disease in a population module; Win Episcope Version 2.0; Thrusfield et al. 2001).
Nineteen of 61 animals (31%; 95% confidence interval [CI], 20%–44%) were positive for antibodies to Leptospira in one or more trapping periods, including 6 of 19 (32%; 95% CI, 13%–57%) juvenile (<1 year) raccoons and 13 of 42 (31%; 95% CI, 18%–47%) adults. Of the 19 positive animals, 14 were seropositive for serogroup Grippotyphosa and 4 were positive for serogroup Pomona antigens. One animal was positive for both serogroups Australis (titer 100) and Grippotyphosa (titer 3200).
The overall seroprevalence of Leptospira rose from 5% (2/21) in June to 38% (9/24) in October (Figure 1). Of nine animals that were observed to develop positive antibody titers from one trapping session to the next, six seroconverted between August and September (Table 2). None of the 10 animals that were serologically negative for Leptospira in September developed antibodies in October. Positive titers ranged from 100 to 6400. Of eight animals that were caught the month after they first developed positive antibody titers, all showed the same or decreased titers the following month (Table 2). One animal was serologically positive with serogroup Pomona for the 4 months it was trapped (including the initial trapping period in June).
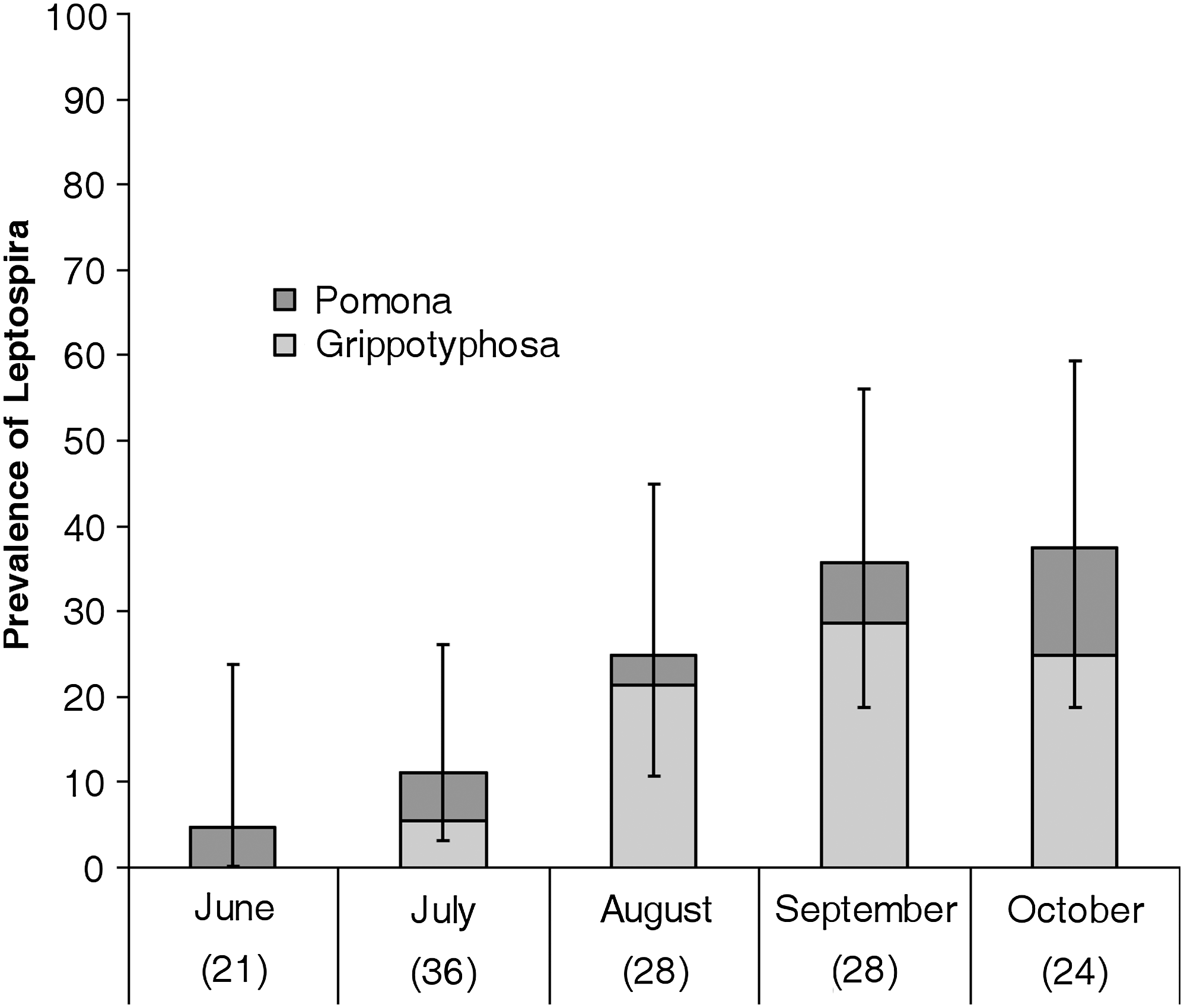
Monthly prevalence of Leptospira serogroups Grippotyphosa and Pomona in raccoons trapped on the grounds of the Toronto Zoo in 2007. Error bars represent 95% confidence intervals. Sample size is shown in brackets below month.
Dashes indicate that the animal was not trapped that session, and 0 indicates that no antibodies were detected. Gray-shaded boxes indicate documented cases of seroconversion from one trapping period to the next.
Also positive for serovar Bratislava (titer 100).
Urine culture positive.
Seven of eight serologically negative animals that were treated with penicillin in June were caught in subsequent months. Of these seven animals, four developed antibodies to Leptospira. Ten of 12 serologically negative animals that were not treated with penicillin in June were sampled in subsequent months, and 3 of these 10 animals developed antibodies to Leptospira in subsequent months.
Urine samples collected from trays under traps were grossly contaminated and unsuitable for culture, and for this reason 35 urine samples were collected by cystocentesis from 23 animals, including 7 animals that were sampled two or more times. Twenty-eight samples were from animals that were serologically negative for Leptospira and seven were from animals with antibodies to Leptospira. Only one sample was culture positive for Leptospira (2.9%). The culture-positive sample was collected in September, at the time the animal had seroconverted (Table 2). This animal was culture negative the following month despite maintaining a positive titer for Leptospira serogroup Grippotyphosa (Table 2). Leptospires grew only in the tube inoculated with 500 μL of the 10−1 dilution after 4 weeks of incubation, so that numbers present were judged to be low.
Discussion
To the best of the authors' knowledge, this is the first study in Ontario that has used repeated trapping of urban raccoons to examine seroconversion for Leptospira and other selected pathogens. We trapped a total of 61 raccoons, including 33 individuals that we were able to capture on multiple occasions over the course of the study.
Raccoons are not thought to play an important role in the ecology of F. tularensis (Morner and Addison 2001), but may act as sentinels for F. tularensis. Our results indicate that the prevalence of F. tularensis during the study period at our site was low. Other researchers have reported low prevalence of antibodies to F. tularensis in raccoon populations [3% in Florida (Bigler et al. 1975)]; however, the majority of studies have reported much higher levels of exposure [38% in Nebraska (Bischof and Rogers 2005); 24.9% in Florida (McKeever et al. 1958)]. Berrada et al. (2006) found that 52% of raccoons were exposed to F. tularensis during a human outbreak of tularemia on Martha's Vineyard, United States, whereas no raccoons trapped on a nearby island (with no outbreak) were serologically positive for F. tularensis.
There was no serological evidence of exposure to avian influenza virus in raccoons in this study. Hall et al. (2008) found that evidence of exposure to avian influenza in raccoons in the United States ranged from 0% to 25% depending on location. The role of raccoons in the ecology of avian influenza is unclear and further studies are required to determine if they are involved in the maintenance or dissemination of influenza virus in nature (Hall et al. 2008).
The overall seroprevalence of leptospiral infection in raccoons in this study was 31%, which is generally comparable to what has been found in raccoons in other studies in a variety of locations [50% in Maryland (Alexander et al. 1972); 58.8% in Georgia (Shotts et al. 1975); 48% in Illinois (Mitchell et al. 1999); 36% in Connecticut (Richardson and Gauthier 2003); 19.1% in Washington (Davis et al. 2008); 47% in Indiana (Raizman et al. 2009)]. Unlike Mitchell et al. (1999) we detected no difference in the seroprevalence of leptospiral infection between adults and juveniles; however, the sample size was relatively small and we cannot rule out the possibility that there was a difference that we were unable to detect.
Penicillin was administered to 9 of 21 raccoons in the initial trapping period as a prophylactic measure following surgical sterilization. Penicillin has been used for the treatment of leptospirosis in humans and animals (Adin and Cowgill 2000, Griffith et al. 2006). No animals had evidence of infection (Leptospires in urine) and only one animal had evidence of exposure to Leptospira (leptospiral antibodies) at the time of treatment, so it was not possible to assess the potential impact of administering penicillin on Leptospira infections in this study.
Antibodies to all seven serogroups tested have been detected in raccoons in previous studies (Mitchell et al. 1999, Raizman et al. 2009); however, there was evidence of exposure to only two serogroups (Grippotyphosa and Pomona) in raccoons in this study. One animal had antibodies to both serogroups Australis and Grippotyphosa, but this dual reaction might be due to cross-reaction of the MAT rather than exposure to serogroup Australis (Levett 2004). The occurrence of antibodies to serogroup Grippotyphosa exhibited a seasonal pattern with the highest prevalence occurring in late summer and early fall. On the basis of the longitudinal data most infection with Leptospira occurred between August and September. This shows that raccoons in this study were at highest risk of being exposed to Leptospira in August and early September.
In the United States and Canada, the highest seroprevalence of Leptospira in clinically affected dogs is typically seen in October and November (Ward 2002, Alton et al. 2009), slightly later than the seroconversion observed in raccoons in this study. Ward (2002) found that there was a significant correlation between the number of cases diagnosed in dogs and the average rainfall recorded 3 months before diagnosis. The slightly earlier onset of Leptospira serogroup Grippotyphosa seen in raccoons in this study could be related to unusual rainfall patterns (less than normal) seen that summer. Alternatively, if the results of multiyear studies show that infection in raccoons consistently occurs before infection in dogs, it might suggest that raccoons have the potential to act as important sources of infection for dogs as suggested by others. The present study finished in October, but it would be interesting to continue into the later months of the fall.
We were only able to isolate Leptospira from the urine of a single animal in this study. Mitchell et al. (1999) found that 6% of urine samples from raccoons in their study were positive for Leptospira. The seroprevalence of Leptospira in their population was higher than that observed in our study (48%), and the higher level of urine shedding that they observed might be due to an increased level of infection in that population (all five culture-positive animals in their study were also seropositive). Leptospira are difficult to culture (Harkin et al. 2003) and it is possible that urine collection and culture techniques were inadequate. Since leptospires were grown only in the medium with the highest inoculum of sample and contamination with other organisms was minimal, greater recovery of leptospires might be achieved with increased inocula, for example direct inoculation into the medium. Using polymerase chain reaction–based assays on kidney and/or urine samples would allow us to avoid these problems in the future; however, current polymerase chain reaction assays do not allow identification of the infecting serovar that is important for epidemiological studies like this one (Levett 2004).
Raccoons have been implicated as the likely reservoir of Leptospira serovar Grippotyphosa in Ontario (Warshawsky et al. 2000, Prescott 2008). We showed that raccoons can shed leptospires in urine, indicating that they may act as a potential source of Leptospira infection for humans and dogs in Ontario. However, there was no evidence that animals were persistently infected with Leptospira, which suggests that they may not be maintaining the pathogen in the environment. One surprising feature of the study was the low seroprevalence of raccoons in the early summer, suggesting that leptospirosis was not endemic in this population. One assumption relating the surge in canine leptospirosis in Ontario observed over the last decade to leptospirosis in raccoons is that leptospirosis in raccoons is common and widespread (Prescott et al. 2002, Alton et al. 2009), and may relate to an observed increase in seroprevalence of raccoons to Grippotyphosa over several years in the United States (Mitchell et al. 1999). This assumption may need revision. It is important to recognize, however, that this was a small scale study that occurred at one site over a short time. Further, large-scale, longitudinal studies investigating the role of the environment and of raccoons and also of other potential wildlife reservoirs are necessary if we hope to unravel the complex ecology of Leptospira in the future.
Footnotes
Acknowledgments
We thank the Toronto Zoo for use of zoo grounds and its excellent staff for assistance in animal capture and handling. We thank Ann Nguyen and Katherine Winger for assistance with trapping and processing animals, and Antonia Dibernardo, Barb Jefferson, and Maria Osuch for their help in the laboratory.
Disclosure Statement
No competing financial interests exist.