
Abstract
Natural sandfly infection by Leishmania spp. in an area endemic for American cutaneous leishmaniasis was analyzed using multiplex polymerase chain reaction (PCR). The sandflies were captured using Falcão light traps in an endemic area of the municipality of Doutor Camargo during March, April, and June 2008. In total, 1803 females were analyzed; 1755 were Nyssomyia neivai (Pinto) and 48 were Nyssomyia whitmani (Antunes and Coutinho). Multiplex PCR analyses using MP3H-MP1L and 5Llcac-3Llcac primers showed the presence of Leishmania (Viannia) spp. in 4/181 pools of sandflies, all Ny. neivai, that is, a minimal infection rate of 0.22%. This study showed, for the first time, the presence of DNA of Leishmania (Viannia) spp. in Ny. neivai. This suggests the existence of natural infection by Leishmania (Viannia) spp. in Ny. neivai in the state of Paraná. Multiplex PCR is an important tool in the detection of Leishmania infection in sandflies.
Introduction
The incidence of American cutaneous leishmaniasis (ACL) has been increasing in Latin America, especially in Brazil, where 634,914 cases were reported from 1980 to 2006. Of these, 14,129 were recorded in the southern region of the country, and 13,762 (97.4%) of these occurred in the state of Paraná (Ministério da Saúde 2009). The high incidence, the wide geographical distribution, and the possibility that ACL will cause destructive, disfiguring, and incapacitating lesions, which can have psychosocial effects on the individual, indicate the importance of this disease (Gontijo and Carvalho 2003).
In Brazil, the transmission cycle of Leishmania spp. involves only a few of the ∼800 species of sandflies that exist worldwide (Aguiar and Medeiros 2003). Studies in northern Paraná show that the sandfly species Nyssomyia whitmani (Antunes and Coutinho), Nyssomyia neivai (Pinto), Pintomyia pessoai (Coutinho and Barretto), Pintomyia fischeri (Pinto), and Migonemyia migonei (França) are always present in the area, and they may be important in the epidemiology of ACL (Teodoro et al. 2001, 2003, 2004, 2007, Membrive et al. 2004, Reinhold-Castro et al. 2008). The detection and identification of the Leishmania species that naturally infect sandflies, as well as the determination of the infection rates of these vectors, are important for understanding their epidemiology and for prevention of the disease in endemic areas (Michalsky et al. 2002, Kato et al. 2005). Luz et al. (2000) reported for the first time in Paraná, the infection of Ny. whitmani by Leishmania (Viannia) braziliensis, demonstrating the need for new studies to detect natural infection in other species of sandflies that are frequently found in endemic areas and that could be involved in the epidemiology of ACL in the state.
In the Recanto Marista, municipality of Doutor Camargo in Paraná, 10 ACL cases were reported in 1996 among the 24 inhabitants of this locality, in addition to nine dogs with positive serology for leishmaniasis (Teodoro et al. 2001). Measures to control the sandfly population were introduced in Recanto Marista; however, these measures have not been rigorously maintained. This probably explains the occurrence of four ACL cases in this locality, one in 2001, one in 2004, and two in 2005 (Reinhold-Castro et al. 2008). The objective of the present study was to investigate natural Leishmania infection in sandflies in an endemic region of Paraná, using the technique of multiplex polymerase chain reaction (PCR).
Materials and Methods
Area of study
The study was carried out in a recreation area named the Recanto Marista, in the municipality of Doutor Camargo (52°13′ W and 23°33′ S), which is located in the North-Central Paraná Mesoregion (Fig. 1). Recanto Marista is located on the banks of the Ivaí River and has a total area of 57.6 ha, of which 40.8 ha are covered by semideciduous seasonal forest, which although heavily impacted is still inhabited by wild mammals. In spite of the deforestation around the Recanto Marista for soybean, maize, and wheat cultivation, the enzootic cycle of Leishmania persists in the remaining forest in this area. Further, peridomiciliar conditions (domestic animal shelters next to the domicile, accumulation of organic substances, wet soil from surface disposal of household wastewater, and shade trees) may favor the formation of sandfly breeding sites and the concentration of these insects in the domicile (Massafera et al. 2005, Monteiro et al. 2008, 2009).
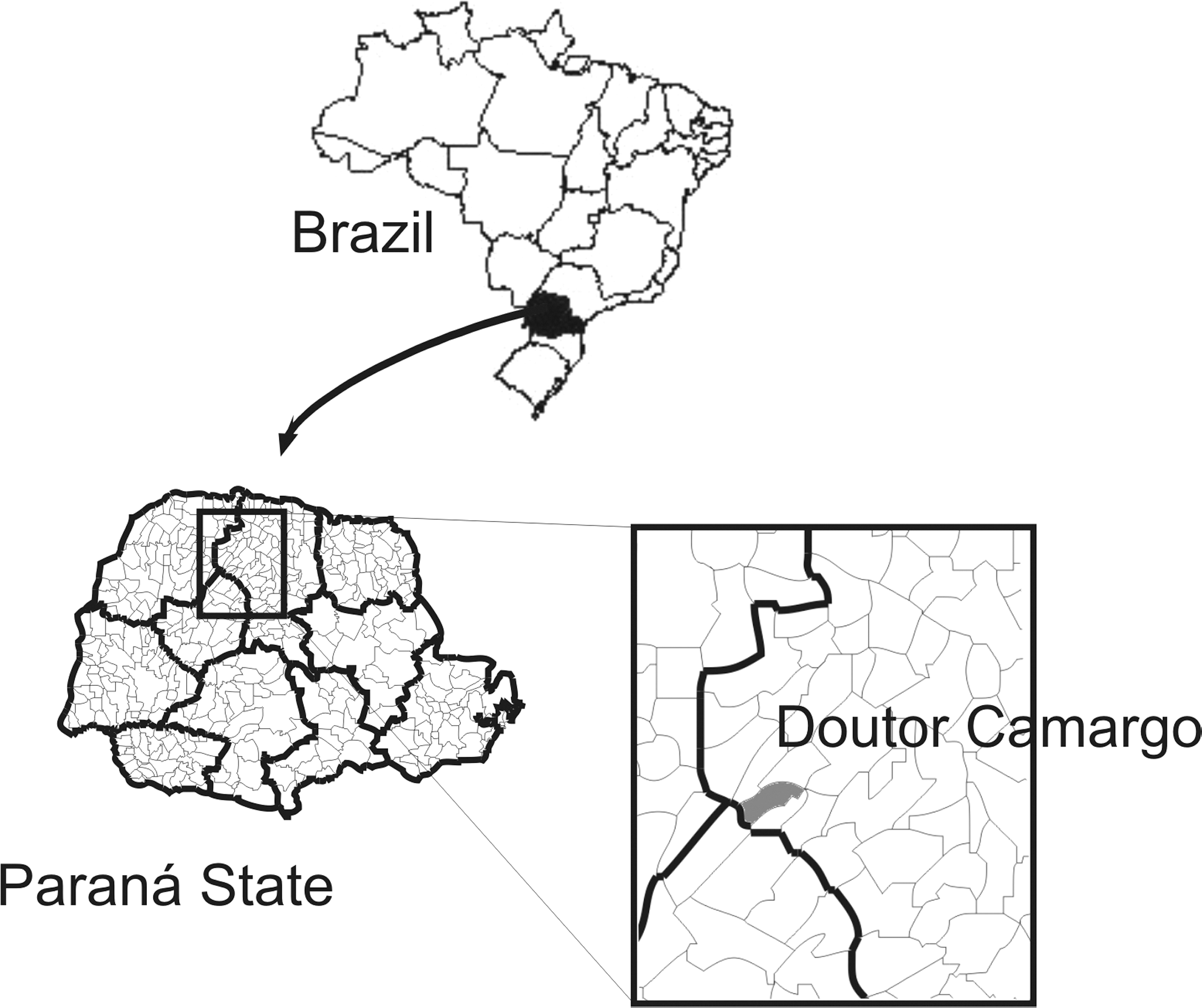
Location of Doutor Camargo municipality in the state of Paraná, Brazil.
Collection of sandflies
Collections were carried out on three different occasions, selected according to the meteorological conditions, especially rainfall, air relative humidity, and air temperature, to increase the chances of collecting as many sandflies as possible. Collections were carried out in March 2008, from 20:00 to 24:00 h; April 2008, from 21:00 to 05:00 h; and July 2008, from 01:00 to 06:00 h. Since the fauna, seasonality, time of highest frequency, and the main sandfly ecotopes were already known (Teodoro et al. 2001, 2003, 2007, Reinhold-Castro et al. 2008), collections were carried out in the ecotopes where the most sandflies were likely to be caught.
The collections were made using Falcão light traps in the following ecotopes: Ecotope 1 (E1): inside a disused hen house next to a house in the forest (collection period: 4 h). Ecotope 2 (E2): on the riverbank next to the forest (at the top of a flight of steps descending to the river) (collection period: 13 h). Ecotope 3 (E3): in the foundation of a house that is occasionally rented to visitors (collection period: 17 h). Ecotope 4 (E4): in the main entrance of a house on the bank of the Ivaí River, used by groups of people as a spiritual retreat (collection period: 9 h).
Identification of the sandflies
The insects were processed at the Entomology Laboratory in the Department of Clinical Analyses of the Universidade Estadual de Maringá, in the following manner: After being killed by chloroform, the insects were placed in tubes containing isopropanol for their preservation and later identification (Paiva et al. 2007). A stereoscopic microscope was used to identify the insects, which were placed on slides washed with 2% sodium hypochlorite and disinfected with 70% ethanol. Two drops of sterile saline (0.9%) were placed on the slides, and the legs and wings of the insect were removed in the first drop, by means of stylets. In the second drop, in the females, two small cuts were made in the final portion of the abdomen to expose the spermatheca, which was placed on slides and examined in an optical microscope, at 200× magnification, to identify the species. The nomenclature of the species followed that of Galati (2003). After identification, the insects were conserved in tubes containing 500 μL isopropanol, each holding 7 to 10 female or 50 male specimens, for later DNA extraction.
Extraction of DNA from the sandflies
To extract the DNA, the insects (pools of 7 to 10 female sandflies) were macerated with a sterile spatula in 20 μL lysis buffer/insect (50 mM NaCl, 10 mM ethylenediaminetetraacetic acid disodium [DTA], pH 8.0, 50 mM Tris-HCl pH 7.4, 1% Triton X-100, and 10 mM dithiothreitol) and were then passed through three cycles of freezing (liquid nitrogen) and defrosting (60°C). The macerate was then incubated for 1 h at 60°C and for 3 more hours at 60°C with the addition of 1 μL proteinase K (20 mg/mL), 80 μL lysis buffer, and 1% Triton X-100 for each insect. After incubation, 300 μL of guanidine isothiocyanate and phenol solution (5 M guanidine isothiocyanate: equilibrated phenol; v/v) was added and vigorously homogenized. Next, 50 μL frozen chloroform was added and gently homogenized. The samples were then centrifuged for 10 min at 9300 g, and the supernatant was transferred to another tube containing 300 μL frozen absolute ethanol. The tube was centrifuged for 15 min at 9300 g, and the sediment was washed twice more with 300 μL frozen absolute ethanol (10 min at 9300 g). The DNA was dried in a dry bath (Bioplus IT-2002) at 95°C, resuspended in 20 μL of distilled H2O, and stored at 4°C until use.
DNA was also extracted from pools of 50 male sandflies. The DNA was quantified by the Qubit™ fluorometer Kit (Invitrogen®) and diluted for the sensitivity tests and the positive control for the reaction.
Culture of Leishmania
Promastigote forms of L. (Viannia) braziliensis (MHOM/BR/1987/M11272) were cultured in medium 199® (Invitrogen) containing 10% bovine fetal serum (Invitrogen), 1% L-glutamine (Invitrogen), and 1% human urine, at 25°C in a B.O.D. incubator (Logen Scientifıc), until the stationary growth phase.
Extraction of Leishmania DNA
The parasites (∼50 mL of culture) were washed four times by centrifugation (10 min at 1600 g) in phosphate buffer solution pH 7.2. The supernatant was discarded, and the DNA was extracted from the pellet by the guanidine-phenol method (Venazzi et al. 2006). The DNA was quantified by the Qubit fluorometer Kit (Invitrogen) and diluted for the sensitivity tests and the positive control for the reaction.
Hot-start multiplex PCR and conventional PCR
Two pairs of primers were used for DNA amplification by multiplex PCR. The MP3H (5′-GAACGGGGTTTCTGTATGC-3′) and MP1L (5′-TACTCCCCGACATGCCTCTG-3′) primers were used to amplify a fragment of 70 bp of the conserved region of DNA from the minicircle of the kinetoplast (kDNA) of the subgenus Leishmania (Viannia) (Lopez et al. 1993). The second pair of primers, 5Llcac (5′-GTGGCCGAACATAATGTTAG-3′) and 3Llcac (5′-CCACGAACAAGTTCAACATC-3′), amplifies a fragment of 220 bp from the IVS6 gene region of the cacophony in insects of the genus Lutzomyia (Lins et al. 2002). The PCR reaction mixture (final volume 25 μL) was composed of 0.5 μM of each of the primers (Invitrogen), 0.2 mM dNTP (Invitrogen), 1 U Platinum Taq DNA Polymerase (Invitrogen), 1.5 mM MgCl2, 1× enzyme buffer, and 2 μL DNA template. The amplification was carried out in a PC Thermocycler (Biometra) at 94°C for 7 min to activate the enzyme (hot-start PCR), followed by 30 cycles, each divided into three stages, of denaturation (1.5 min at 95°C), annealing (1.5 min at 57°C), and polymerization (2 min at 72°C). After this, the extension was continued for a further 10 min at 72°C, and the tubes were then kept at 4°C until analysis.
For the conventional PCR, the primers B1 (5′-GGGGTTGGTGTAATATAGTGG-3′) and B2 (5′-CTAATTGTGCACGGGGAGG-3′), which amplify a fragment of 750 bp of the kDNA minicircle region of species of the L. braziliensis complex (De Bruijn and Baker 1992), were used. The reaction mixture (total volume 25 μL) was composed of 1 μM of each of the primers (Invitrogen), 0.2 mM dNTP (Invitrogen), 1 U Taq DNA Polymerase (Invitrogen), 1.5 mM MgCl2, 1× enzyme buffer, and 2 μL DNA template. The DNA amplification was carried out at an initial temperature of 95°C for 5 min, followed by 35 cycles, each divided into three stages, of denaturation (1.5 min at 95°C), annealing (1.5 min at 60.5°C), and polymerization (2 min at 72°C), and the final extension for 10 min at 72°C. The tubes were kept at 4°C until analysis.
The amplification products were submitted to electrophoresis in agarose gel (2% for multiplex PCR and 1% for conventional PCR), stained with 0.1 μg/mL ethidium bromide, at 10–15 V/cm. For every five samples, one positive and one negative control were added. The presence of bands was observed in a transilluminator (Hoefer MacroVue UV-20®) at 302 nm.
PCR sensitivity assay
PCR sensitivity was determined with the 5Llcac-3Llcac primers to analyze the serial dilutions of the DNA of the male sandflies. DNA template from L. braziliensis promastigotes (MHOM/BR/1987/M11272), serially diluted, was used for the analysis of the sensitivity of the PCR carried out with the MP3H-MP1L and B1-B2 primers.
To determine the sensitivity of the multiplex technique for detecting Leishmania DNA, different concentrations (64, 32, 16, 8, 4, 2, 1 fg) of DNA from L. braziliensis were added to 384 pg of male sandfly DNA and analyzed.
Minimal infection rate
The minimal infection rate was calculated as: minimal infection rate = number of positive pools × 100/total number of sandflies analyzed (Paiva et al. 2007).
Results
In total, 1803 samples of female sandflies were analyzed, of which 1755 (97.3%) were Ny. neivai (Pinto) and 48 (2.7%) were Ny. whitmani (Antunes and Coutinho). Table 1 shows the ecotopes in which the sandflies were captured.
E1: inside a disused hen house next to a house in the forest; E2: on the riverbank next to the forest (at the top of a flight of steps that descends to the river); E3: in the foundations of a house occasionally rented to visitors; E4: in the main entrance of a house on the bank of the Ivaí River, used by groups of people as a spiritual retreat.
The analytic sensitivities of the MP3H-MP1L and 5Llcac-3Llcac primers, individually tested by conventional PCR, were 2 fg of L. braziliensis DNA (Fig. 2A) and 12 pg of Ny. neivai DNA (Fig. 2B), respectively. However, when tested together by multiplex PCR, the detection limit was 16 fg of L. braziliensis DNA (Fig. 2C). The B1-B2 primer pair demonstrated a detection limit of 4 fg of L. braziliensis DNA using conventional PCR (Fig. 2D).

Representative gel showing analytic sensitivity assays of the (
Multiplex PCR analyses were carried out for all the female sandflies captured. The 220-bp fragment was detected in all the sandfly DNA samples tested, demonstrating the absence of Taq DNA polymerase inhibitors.
The results obtained from the multiplex PCR showed the presence of Leishmania (Viannia) spp. in 4/181 pools of the sandflies tested (Fig. 3A). These results were confirmed by conventional PCR using the B1-B2 primers (Fig. 3B). The minimal infection rate, assuming one sandfly was infected in each positive pool, was 0.22% or 4/1803 females analyzed. Since all the positive pools consisted of sandflies from the species Ny. neivai, the minimal infection rate of Ny. neivai was 0.23% (4/1755).
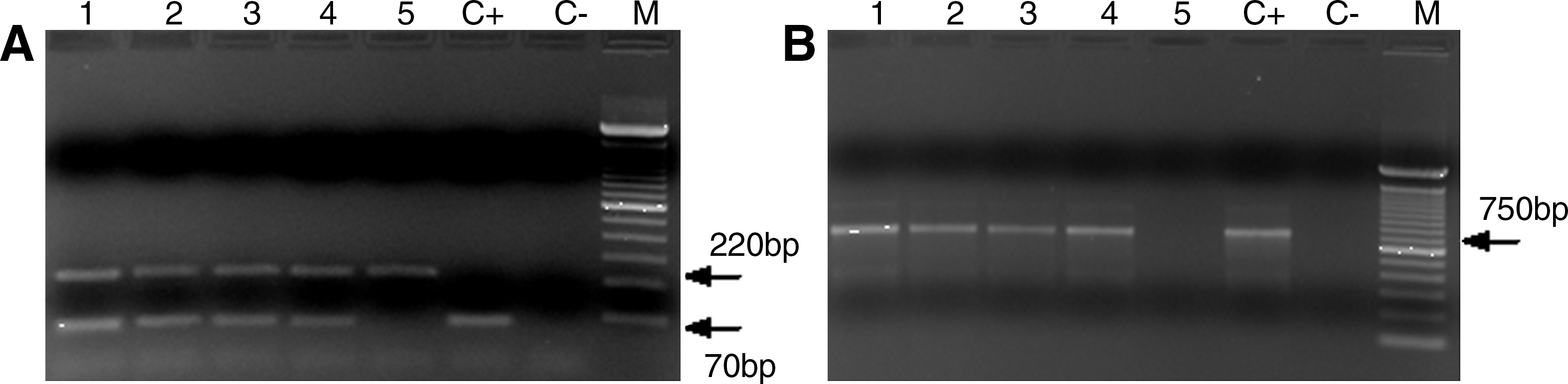
Representative gel showing (
Discussion
In the majority of studies on the natural infection of sandflies by Leishmania, the insects first needed to be dissected to be subsequently analyzed by PCR. This procedure must be carried out soon after collection, preferably in the first few hours, due to the insect's low resistance outside its natural environment and the inviability of carrying out the procedure after it has been dead for some time. In the present study, however, dissection was not carried out for the observation of natural infection by Leishmania in the sandflies. The insects collected were conserved in isopropanol (Paiva et al. 2007), which permitted the collection of a large number of specimens, enabling later processing of the insect and the preservation of its DNA.
For DNA extraction, only the legs and wings were removed. Paiva et al. (2007) demonstrated that the results are better when the whole insect is used for DNA extraction and not just the digestive tract. To validate the protocol used for the extraction of DNA from the sandfly, the quantity of DNA obtained and the absence of Taq DNA polymerase inhibitors were taken into consideration and confirmed by the results obtained in the multiplex PCR.
The use of PCR to determine Leishmania infection in sandflies captured in the field is a useful procedure when there is a large number of samples, enabling the identification of suspected vectors (Perez et al. 1994, Rodríguez et al. 1999). According to Feliciangeli et al. (1988), PCR is more specific and sensitive than dissection for the determination of infection in sandflies. The sensitivity obtained with conventional PCR using the MP3H-MP1L primers was 2 fg, similar to that found by Lopes et al. (1993) and Velásquez et al. (2006), who detected 0.14 and 0.9 fg of DNA, respectively. The sensitivity using the B1-B2 primers was 4 fg of DNA, also similar to the 1 fg reported by De Bruijn and Baker (1992).
To be sure that the negative results were valid, and not due to inhibition of the Taq DNA polymerase by other possible inhibitor substances present in the insects, a multiplex PCR assay associating the MP3H-MP1L primers with the 5Llcac-3Llcac primers, which amplify the gene of the cacophony in Lutzomyia (internal amplification control), was standardized. This gene has been used as an important internal control for the activity of Taq DNA polymerase in PCR for Lutzomyia (Pita-Pereira et al. 2005).
Although the multiplex PCR assay showed lower analytical sensitivity than that of conventional PCR with the MP3H-MP1L primers, it was still very sensitive for the detection of DNA from the subgenus Leishmania (Viannia). Further, the use of primers that perform the function of an internal control for the reaction gives greater assurance that the results are valid, because the insects have in their tissues, especially in their exoskeleton, head, and thorax, inhibitors that can reduce the efficiency of the PCR reaction (Higgins and Azard 1995, Siridewa et al. 1996). The positive results obtained in the multiplex PCR, which showed the presence of DNA from Leishmania (Viannia) spp. in 4/181 pools of sandflies (2.2%), were confirmed by the conventional PCR with the B1-B2 primers, which amplify DNA from species of the L. braziliensis complex.
Assuming that one sandfly was infected in each pool, the infection rate found in this study was 0.22% (4/1803). Note that all the positive pools were of the species Ny. neivai. Then the minimal infection rate of Ny. neivai was 0.23% (4/1755). Although ACL is endemic in Paraná, infection in sandflies has been little studied in this state. The first reported isolation and identification of L. braziliensis in sandflies in the state was carried out by Luz et al. (2000), who found three Ny. whitmani females naturally infected by L. braziliensis out of a total of 1612 dissected female sandflies (0.18%) in the municipality of Cambira in northern Paraná. The prevalence of Ny. whitmani in some municipalities and of Ny. neivai in others was previously reported in northern Paraná (Membrive et al. 2004, Teodoro et al. 2006). In the municipality of Adrianópolis, also in Paraná, Castro et al. (2005) did not detect any Leishmania infection in 2559 dissected females of the species Nyssomyia intermedia (2530), Pi. fischeri (15), Mi. migonei (11), and Ny. whitmani (3).
PCR has been widely used in the study of the natural or experimental infection of sandflies, using various primers (Pérez et al. 1994, Rodriguez et al. 1999, Aransay et al. 2000, Michalsky et al. 2002, Miranda et al. 2002, Jorquera et al. 2005, Kato et al. 2005, Pita-Pereira et al. 2005). In Paraná, using dissection and conventional PCR, Neitzke et al. (2008) analyzed 2487 females and Scodro et al. (2008) analyzed 2213 females, the majority of which were Ny. neivai, but they did not detect any infection. In the state of Rio de Janeiro, Pita-Pereira et al. (2005) analyzed 40 pools of females by multiplex PCR and hybridization and found that 5/32 Ny. intermedia and 3/5 Mi. migonei were infected with L. braziliensis, that is, an infection rate of 2% for the insects. In the state of Rio Grande do Sul, Pita-Pereira et al. (2009) analyzed 27 pools of Ny. neivai females by multiplex PCR and found an infection rate of 1.1%.
Although the detection of infection by PCR does not fulfill the requirements of vector incrimination proposed by Killick-Kendrick and Ward (1981), experimental infections of Ny. intermedia (Silva and Gomes 2001) have shown that this species is susceptible to the strains of L. braziliensis and have demonstrated their locations in the digestive tract. This information, together with the epidemiology of ACL, indicates that the sandflies of the Ny. intermedia s.l. complex may play an important role in the epidemiology of ACL (Marcondes 1996). Natural infection of Ny. neivai has been reported by Marcondes et al. (2009) in Santa Catarina state, Brazil, using the techniques of PCR and Southern blot hybridization, and by Córdoba-Lanús et al. (2006) in northwest Argentina using PCR and hybridization.
Ny. neivai was predominant in the various studies that have been carried out in the municipality of Recanto Marista (Teodoro et al. 2001, 2003, 2007, Reinhold-Castro et al. 2008). We had intended to assess the infection rates in the five species of sandflies that are most frequently found in this locality. However, the number of specimens of other species collected was insufficient to make a pool (only one Pi. fischeri, one Pi. pessoai, and no Mi. migonei were collected). The infected sandflies were captured in the E2 (river bank), E3 (house foundations), and E4 (house) ecotopes; the last two locations are periodically frequented by visitors. The E2 and E4 ecotopes are close to a forest bordering a river, and the E3 ecotope is ∼150 m from this forest, where small wild mammals live that could be natural reservoirs of Leishmania spp. This could explain the existence of naturally infected sandflies. This information, together with other reports of natural infection of this sandfly species in other endemic regions, confirms the importance of Ny. neivai in the epidemiology of ACL. Further, they show the importance of controlling the insect population to avoid ACL in the area studied, because several autochthonous cases of ACL have already occurred in this locale.
This is the first report of DNA from Leishmania (Viannia) spp. in Ny. neivai in the state of Paraná. In Paraná, the parasites isolated from human and canine ACL cases have been identified as L. braziliensis (Lonardoni et al. 1993, Silveira et al. 1999). This suggests that the specimens of Ny. neivai were naturally infected with L. braziliensis.
This study shows that multiplex PCR containing an internal amplification control is a useful tool in the detection of Leishmania infection in sandflies. The detection of DNA from Leishmania (Viannia) spp. in Ny. neivai indicates that this sandfly species may play an important role in the transmission of L. braziliensis in the state of Paraná.
Footnotes
Acknowledgments
The authors thank the Fundação Araucária and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq—award number 410550/2006-0) for financial support.
Disclosure Statement
No competing financial interests exist.