
Abstract
Rift Valley fever virus continues to cause large outbreaks of acute and febrile illness among humans and domestic animals in Africa. The high pathogenicity of the virus is mainly due to the non-structural protein derived from the S segment NSs, which was shown to inhibit the type I interferon expression at the transcriptional level and to suppress host cell RNA synthesis. Clone 13, a naturally attenuated clone containing a deletion of 70% in NSs, is a promising vaccine candidate as it has no pathogenicity for mice and is highly immunogenic leading to long-lasting immunity. If Clone 13 succeeds in inducing a transient viremia in inoculated animals, is a mosquito vector able to replicate Clone 13 and is the vector affected by viral infection? In this work, we orally infected two mosquito species, Aedes vexans and Culex pipiens quinquefasciatus, with the avirulent Clone 13. We showed that the mosquito Ae. vexans better replicated the avirulent Clone 13 than Cx. p. quinquefasciatus. Moreover, infection with Clone 13 did not cause any important changes in mosquito's life-history traits compared to noninfected females. Nevertheless, it is likely that Clone 13 would not be efficiently transmitted by mosquito vectors.
Introduction
Numerous species of arthropods have been incriminated as vectors. Different mosquito species are involved in various geographic areas where the disease is endemic; more than 30 species have been found naturally infected by RVFV, mostly the Aedes and Culex genera. RVFV could be maintained in an enzootic cycle involving Aedes mosquitoes such as Ae. vexans in West Africa (Fontenille et al. 1995, 1998, Zeller et al. 1997, Traore-Laminaza et al. 2001). Aedes species are able to transmit the virus vertically to their offspring (Linthicum et al. 1985, 1999). Epizootic outbreaks are often linked with unusual rains or warm seasons, favoring the hatching of infected Aedes eggs that are then able to initiate the virus circulation. Subsequently, large numbers of secondary vectors could be infected and induce the emergence of epidemic/epizootic outbreaks. Culex mosquitoes are abundant in peri-urban areas. For example, Cx. pipiens pipiens was implicated in Egypt and the Arabian Peninsula (Meegan et al. 1980, Jupp et al. 2002, Miller et al. 2002), whereas Cx. pipiens quinquefasciatus is present in high densities during RVF outbreaks and has been suspected to play a role in mosquito–human transmission (Marrama et al. 2005). The emergence of RVF usually corresponds to the conversion from enzootic to epizootic virus activity, which follows heavy rains leading to flooding in grassland zones in Africa. Aedes species ensure the emergence of the virus acquired through vertical transmission, and Culex species ensure the amplification and dissemination of the virus out of the enzootic/endemic zones. Moreover, it is known that transmission to humans occurs through direct contacts with high virus loads when aborted fetuses are manipulated.
RVFV (Bunyaviridae family, Phlebovirus genus) is a tri-segmented negative-stranded RNA virus composed of three segments: the L segment, coding for the RNA polymerase RNA dependent; the M segment, coding for the glycoprotein precursor GN and GC; and the S segment, which has an ambisense strategy, coding for the nucleoprotein N and the non-structural protein derived from the S segment NSs (Giorgi et al. 1991). The phosphoprotein NSs plays a key role in the pathogenesis of RVFV (Bridgen et al. 2001, Weber et al. 2002). This nonstructural protein antagonizes the antiviral response by blocking type I interferon (IFN) production (Bridgen and Elliott, 1996). The protein NSs appears to be nonessential for the RVFV cycle since it exists in a natural virus Clone 13, in which 70% of the NSs Open reading frame (ORF) is deleted (Muller et al. 1995, Vialat et al. 2000).
Clone 13, an isolate biologically cloned by plaque purification from a nonfatal human case during the 1974 outbreak in Bangui, Central African Republic (Muller et al. 1995), presents a large deletion in the NSs segment. The major role played by IFN was demonstrated by inoculating mice lacking the alpha/beta interferon receptor with Clone 13: these IFN-deficient animals died very rapidly after inoculation with Clone 13 (Bouloy et al. 2001). Clone 13 is not pathogenic for normal mice or hamsters; animals survive after exposure to large infectious doses of ≤106 plaque-forming units (PFUs) without developing any signs of disease. Moreover, Clone 13 is highly immunogenic, leading to long-lasting immunity (Muller et al. 1995). These particular characteristics have led to propose Clone 13 as a vaccine candidate. It has been tested in sheep and cattle, and all vaccinated animals elicit immune responses protective against challenge with virulent RVFV (Hunter, 2001). If Clone 13 succeeds to induce a transient viremia in inoculated animals as stated by Muller et al. (1995), is a mosquito vector able to replicate Clone 13 and is the vector's life affected by viral infection? In the present study, we demonstrated that, compared to Cx. p. quinquefasciatus, Ae. vexans ensured a better viral replication of Clone 13. Moreover, females of both species infected with Clone 13 did not show any significant changes in their life-history traits compared to noninfected females.
Materials and Methods
Mosquitoes
Two laboratory colonies established for over 10 years were used: (1) Ae. vexans belonging to the Ae. vexans complex and originating from Germany; it has been adapted to laboratory conditions by R. Kuhn (University of Mayence) and is the only Ae. vexans colony available in Europe; and (2) Cx. p. quinquefasciatus (SLAB strain) originated from California (Georghiou et al. 1966). Ae. vexans could be viewed as a species incriminated in the RVFV enzootic cycle and Cx. p. quinquefasciatus as a species involved in the epizootic/epidemic cycle.
Mosquitoes were maintained in insectaries at 28°C ± 1°C with 80% relative humidity and a light:dark cycle of 16 h:8 h. Larvae were reared individually until adult stage in 12-well culture plates containing 4 mL of dechlorinated tap water supplemented with 0.01 g of fish food (Tetramin Baby). Emerging adults were maintained on a diet of 10% sucrose solution before being exposed to virus.
Viruses
The avirulent strain Clone 13 was cloned by plaque purification from the 74HB59 RVF strain isolated in 1974 in Bangui (Central African Republic) from a nonfatal human case (Muller et al. 1995). For our experiments, a monolayer of Ae. albopictus C6/36 cells was infected with the viral stock of Clone 13 produced in Vero cells, at a multiplicity of infection of 0.1 PFU/cell and incubated at 27°C on Leibovitz (L-15) medium (Gibco/Invitrogen) supplemented with 10% fetal bovine serum, 1000 units/mL penicillin, 1 mg/mL streptomycin, and tryptose phosphate broth 1×. Cell infection was checked by indirect immunofluorescence assays using ascetic fluids (RVF Ascite from mouse, and fluorescein isothiocyanate mouse from goat). When 80% of cells were infected (usually 72 h postinfection [pi]), the supernatant fluid was collected and viral titer was estimated by serial 10-fold dilutions on Vero cells (Billecocq et al. 1996). As a control for RVFV virulence, we used the strain ZH548 isolated in 1977 from a human case in Egypt (Meegan 1979). Stocks of ZH548 were produced in Vero cells as described previously for Clone 13.
Infection of cells
African green monkey kidney (Vero) cells were maintained at 37°C with 5% CO2 in Dulbecco's modified Eagle's medium containing 5% fetal bovine serum, 1000 units/mL penicillin, 1 mg/mL streptomycin, tryptose phosphate broth 1×, and HEPES 0.01 M. Ae. albopictus C6/36 cells were maintained at 27°C as described previously. A monolayer of cells (C6/36 or Vero cells) was infected with the stock of Clone 13 at a multiplicity of infection of 5 PFU/cell. Virus was adsorbed to confluent cell monolayers for 1 h at 28°C for C6/36 and 37°C for Vero. At the end of 1 h absorption period, the inoculum was removed and 5 mL of the medium was added. Supernatants were harvested at different hours after infection (2, 4, 6, 8, 12, 16, 20, 24, and 32). Infectious viral particles were titrated by plaque assay (Billecocq et al. 1996).
Infection of mosquitoes
One-week-old female mosquitoes were starved 24 h before infection, and 15–20 batches of 70 females were blood fed. The mixture with two-thirds washed sheep erythrocytes, one-third virus suspension, and adenosine triphosphate added as phagostimulant (5 × 10−3 M) gave a blood meal at a titer of 108.5 PFU/mL, which is the minimal titer required to initiate RVFV disseminated infection (Moutailler et al. 2007). The blood meal was provided on a cotton stick support as described in Moutailler et al. (2007). After a 30 min blood feeding, fully engorged females were sorted on ice and individually isolated in 50 mL plastic tubes. To allow Cx. p. quinquefasciatus females to oviposite, 4 mL of water was dropped inside the tube with a syringe 4 days after blood feeding. For Ae. vexans females, a moist filter paper was provided as substrate to oviposition. Females were supplied with a 10% sucrose solution in a cotton pad put on the mesh covering the tube. Two trials were performed for life-history trait studies and one for the quantitative reverse transcription (RT)-polymerase chain reaction (PCR) analysis. The infection status was checked by immunofluorescence assay on head squashes (Kuberski and Rosen 1977). To estimate the disseminated infection rate, females were kept for 14 days at 28°C. For studies on life-history traits, females were kept until death at 28°C. Disseminated infection rate corresponds to the proportion of females positive by immunofluorescence assays on head squashes among surviving females 14 day pi.
Female life-history traits
Three traits were examined: survival, oviposition, and egg hatching. Mosquito mortality was scored daily to estimate female life span (the number of days from exposure to an infectious blood meal until death). Oviposition was examined through three parameters describing fecundity: (1) the time to first laying to estimate the length of gonotrophic cycle, (2) the number of laid eggs per female, and (3) the time between first oviposition and mosquito death. Hatching was studied by estimating (1) the hatching capacity corresponding to the proportion of mosquitoes with at least one egg hatched among laid eggs, and (2) the hatching rate referring to the proportion of eggs hatched among those laid per mosquito. These two parameters described female fertility. Eggs were stored at 28°C and put to hatch by immersion in dechlorinated tap water.
Quantitative RT-PCR analysis
Three mosquitoes were killed every day and, if existing, dead mosquitoes were collected daily. Mosquitoes were examined until day 14 pi. For each mosquito, total RNA was extracted using the Nucleospin RNA II kit (Macherey-Nagel) following the manufacturer's instructions. Briefly, individual mosquito was ground in 350 μL of lysis buffer and 3.5 μL of β-mercaptoethanol. The lysate was then filtered through filter units and centrifuged for 1 min at 11,000 g. The filtrate collected in a tube was mixed with 70% ethanol. The solution was passed through a column that binds RNA after centrifugation for 30 s at 8000 g. After desalting the silica membrane (centrifugation at 11,000 g for 1 min), a DNAse reaction mixture was applied on the silica membrane of the column for 15 min at room temperature. After different cycles of washing, the RNA solution was eluted by centrifugation at 11,000 g for 1 min in 40 μL of RNAse-free H2O.
To build the standard curve, an RVFV RNA synthetic transcript was generated. A PCR product encompassing the targeted region was prepared using the M segment of ZH548 RVFV strain and cloned into pGem 4Z vector (Promega). The amplified product using vector-specific primers was purified using the PCR Purification Kit (Qiagen). RNA transcripts were produced in vitro using the RiboMAX™ Large Scale RNA Production Systems (Promega) appropriate for either SP6 or T7 RNA polymerase. The transcript size was 3603 bp for RVFV. Residual DNA was eliminated by several DNAse treatments (Turbo DNA-free; Ambion). After quantification by spectrophotometer, RNA transcript solution was stored at −80°C.
The one-step RT-PCR was performed in a volume of 25 μL containing 3 μL RNA template, 12.5 μL 2 × Brilliant SYBR Green I quantitative PCR Master Mix (Stratagene), 0.32 μL sense (50 ng/μL), 0.36 μL antisense (50 ng/μL), 0.25 mL Fluorescein (1 μM), and 0.0625 μL Stratascript RT/RNAse block enzyme. Primers were selected in the M segment of RVFV: sense, 5′-CATGGATTGGTTGTCCGATCA-3′; antisense, 5′-TGAGTGTAATCTCGGTGGAAGGA-3′. The amplification program in an i-Cycler™ (Biorad) included a reverse transcription at 50°C for 30 min, an inactivation step of RT/RNAse enzyme at 95°C 10 min, followed by 40 cycles of 95°C 30 s, 57°C 1 min, 72°C 30 s, a step at 95°C 1 min, and 81 cycles of 55°C (+0.5°C/cycle) 30 s. The size of the amplification product was 108 bp. After amplification, a melting curve was acquired to check the specificity of PCR products. PCR was performed in triplicate for each mosquito. Signals were normalized to the standard curve using serial dilutions of RNA synthetic transcripts. Normalized data were used to measure the number of RNA copies in infected mosquitoes according to the ΔC t analysis.
Statistical analysis
Disseminated infection rates were compared using a χ 2-test. Survival rates after exposure to an infectious blood meal were described using Kaplan–Meier survival curves, which were compared with the logrank test. To assess any effects of an infectious blood meal on female oviposition, the time to first egg laying was estimated for each mosquito and compared using Kaplan–Meier curves and the logrank test. The total number of eggs laid per mosquito was investigated using a negative binomial regression model, which provides incidence rate ratios (IRRs) and their 95% confidence intervals (CIs). The significance level of the covariate was tested using Wald's test. We also analyzed the time between first oviposition and mosquito death using Kaplan–Meier curves and the logrank test. To estimate any changes in egg hatching after exposure of females to an infectious blood meal, the hatching capacities were compared using a Fisher's exact test and the hatching rates using an analysis of variance. All statistical analyses were performed using STATA software (StataCorp LP, Texas, USA).
Results
Replication in vitro
Replication of Clone 13 in C6/36 cells started to increase at 6 h pi and became steady at 24 h pi (Fig. 1). When cultures of Vero cells were infected with Clone 13, the virus production increased at 8 h pi and reached a plateau from 12 h pi. Both cell types ensure a high viral production going up to 109 PFU/mL. Nevertheless, viral replication in Vero cells was faster ending by a lysis of cells from 32 h pi. On the other hand, C6/36 cells remained unaffected by viral replication.
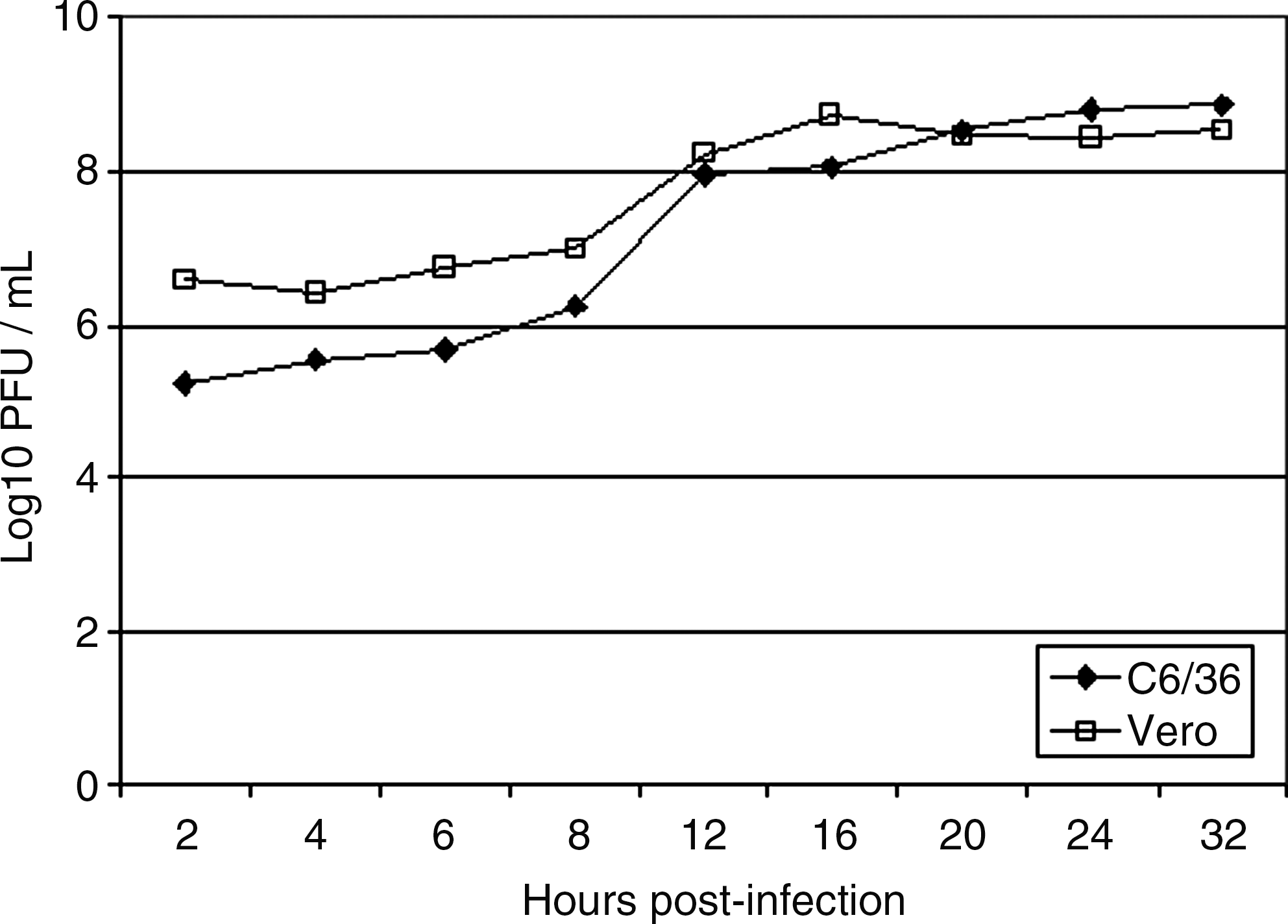
Replication of Clone 13 in C6/36 and Vero cells after infection at a multiplicity of infection of 5 plaque-forming unit (PFU)/mL.
Disseminated infection rates
We showed that for each mosquito species, no significant difference in disseminated infection rates was observed between the two trials (χ 2-test: p = 0.29 and p = 0.26 for Ae. vexans and Cx. p. quinquefasciatus, respectively). Moreover, Ae. vexans mosquitoes showed significantly higher disseminated infection rates than Cx. p. quinquefasciatus (χ 2-test: p < 10−4 and p = 0.002 for trial 1 and trial 2, respectively): 29.3% (trial 1) and 24.7% (trial 2) for Ae. vexans, and 10.2% (trial 1) and 13.7% (trial 2) for Cx. p. quinquefasciatus (Table 1). In addition, Ae. vexans exhibited similar disseminated infection rates when infected with Clone 13 and with the virulent strain ZH548 (χ 2-test: p = 0.44). However, Cx. p. quinquefasciatus presented a higher disseminated infection rate when infected with the virulent strain ZH548 rather than with Clone 13 (χ 2-test: p = 0.03).
N, number of females analyzed; SD, standard deviation; NI, non-infected.
Life-history traits
Survival
Table 1 and Figure 2 indicate respective life spans and survival curves of Ae. vexans and Cx. p. quinquefasciatus after ingestion of an infectious blood meal containing Clone 13 or ZH548. Aedes vexans survived significantly longer when noninfected or infected with Clone 13 rather than when infected with ZH548 (log-rank test: p = 0.0003): 21.2 ± 9.6 (mean no. of days ± standard deviation [SD]) (trial 1) and 21.8 ± 8.2 (trial 2) when infected with Clone 13, 18.9 ± 8.8 when infected with the virulent strain ZH548, and 20.4 ± 9.4 when noninfected. Besides, Cx. p. quinquefasciatus survived differently whatever the infection status: 13.4 ± 7.1 (mean no. of days ± SD) (trial 1) and 14.6 ± 5.9 (trial 2) when infected with Clone 13, 14.9 ± 8.0 when infected with ZH548, and 15.8 ± 7.0 when noninfected. Moreover, Ae. vexans survived significantly longer than Cx. p. quinquefasciatus.
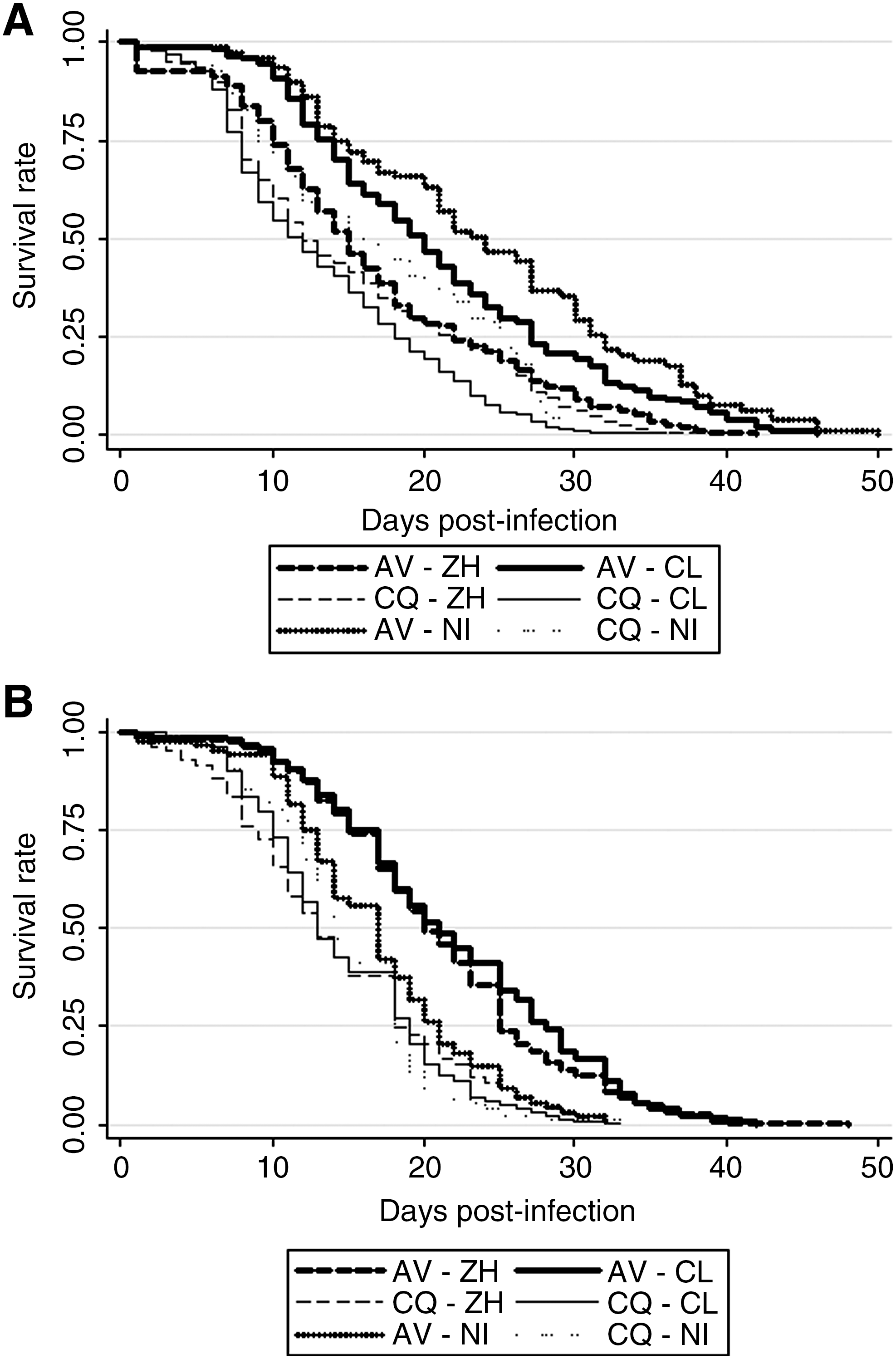
Survival rates of Aedes vexans and Culex quinquefasciatus after an infectious blood meal with Clone 13 (cL) or ZH548 (ZH), or after a non-infectious blood meal (NI). Two trials were carried out (
Oviposition
Three parameters described the oviposition were the time to first egg laying, the number of eggs laid per mosquito, and the time between first oviposition and mosquito death (Table 1).
Time to first laying: Ae. vexans took significantly more time for the first egg laying when noninfected or infected with Clone 13 than when infected with ZH548 (log-rank test: p = 0.003): 19.2 ± 9.9 (mean no. of days ± SD) (trial 1) and 20.6 ± 8.8 (trial 2) when infected with Clone 13, 17.8 ± 9.1 when infected with ZH548, and 19.3 ± 9.8 when noninfected. For Cx. p. quinquefasciatus, the same pattern was obtained whatever the infection status: infected with Clone 13 (7.0 ± 4.4 [mean no. days ± SD], trial 1; 8.7 ± 4.8, trial 2), infected with ZH548 (7.0 ± 4.1), or noninfected (7.9 ± 4.8). In addition, Ae. vexans took significantly more time for the first egg laying than Cx. p. quinquefasciatus.
Number of eggs laid: Using a negative binomial regression model, we found that Ae. vexans infected with Clone 13 laid slightly fewer eggs than noninfected females or those infected with ZH548 (IRR [95% CI] of 0.75 [0.58–0.97]): 19.1 ± 15.4 (mean no. of days ± SD) (trial 1) and 17.2 ± 21.0 (trial 2) when infected with Clone 13, 20.6 ± 19.1 when infected with ZH548, and 20.6 ± 17.5 when noninfected. Cx. p. quinquefasciatus infected with Clone 13 laid a quite similar number of eggs like noninfected females or those infected with ZH548 (IRR [95% CI] of 1.15 [1.02–1.30]): 31.0 ± 11.3 (mean no. of days ± SD) (trial 1) and 36.0 ± 15.5 (trial 2) when infected with Clone 13, 34.1 ± 11.9 when infected with ZH548, and 34.7 ± 12.6 when noninfected. Moreover, Ae. vexans laid less eggs than Cx. p. quinquefasciatus.
Time between first oviposition and female death: Ae. vexans did not behave differently when infected or noninfected (log-rank test: p = 0.05): 5.9 ± 8.3 (mean no. of days ± SD) (trial 1) and 3.4 ± 6.1 (trial 2) when infected with Clone 13, 3.3 ± 5.5 when infected with ZH548, and 2.9 ± 5.2 when noninfected. Cx. p. quinquefasciatus had significantly shortened time between female oviposition and death when infected with Clone 13 compared to females infected with ZH548 or noninfected females (log-rank test: p = 0.0001): 8.5 ± 6.9 (mean no. of days ± SD) (trial 1) and 7.4 ± 6.3 (trial 2) when infected with Clone 13, 9.6 ± 7.9 when infected with ZH548, and 10.7 ± 7.2 when noninfected. In addition, Ae. vexans showed a shorter time between female oviposition and death than Cx. p. quinquefasciatus.
Hatching
Two parameters were used to estimate hatching: the hatching capacity and the hatching rate per female (Table 1).
Hatching capacity: When comparing hatching capacities, Ae. vexans did not show any difference according to the female infection status (Fisher's exact test: p = 0.50): 0.97 ± 0.18 (mean% ± SD) (trial 1) and 0.93 ± 0.25 (trial 2) when infected with Clone 13, 0.95 ± 0.20 when infected with ZH548, and 0.93 ± 0.25 when noninfected. Besides, Cx. p. quinquefasciatus infected with Clone 13 showed significantly lower hatching capacities than females infected with ZH548 or noninfected females (Fisher's exact test: p = 0.01): 0.71 ± 0.45 (mean% ± SD) (trial 1) and 0.73 ± 0.45 (trial 2) when infected with Clone 13, 0.79 ± 0.40 when infected with ZH548, and 0.81 ± 0.39 when noninfected.
Hatching rate per female: Ae. vexans did not show any difference according to the female infection status (ANOVA: p = 0.91): 0.74 ± 0.36 (mean no. of eggs hatched ± SD) (trial 1) and 0.69 ± 0.38 (trial 2) when infected with Clone 13, 0.71 ± 0.37 when infected with ZH548, and 0.69 ± 0.40 when noninfected. In addition, Cx. p. quinquefasciatus presented significantly lower hatching rates when infected with Clone 13 (ANOVA: p = 0.001): 0.16 ± 0.23 (mean no. of eggs hatched± SD) (trial 1) and 0.28 ± 0.32 (trial 2) when infected with Clone 13, 0.31 ± 0.32 when infected with ZH548, and 0.26 ± 0.30 when noninfected. Moreover, eggs of Ae. vexans usually tended to hatch more successfully than eggs of Cx. p. quinquefasciatus.
Replication of Clone 13 in mosquitoes
Each Ae. vexans ingested an average of 105.9 ± 100.02 (mean RNA copies ± SD) at day 0 in the blood meal. After a decrease in viral load from day 1 to 4 pi, most females harbored more than 106 RNA copies (Fig. 3A). However, for some females, the number of RNA copies during the experiment was lower than at day 0, indicating that these individuals did not replicate the viral genome. Culex. p. quinquefasciatus ingested an average of 104.7 ± 100.06 (mean RNA copies ± SD) during the blood meal (day 0). The estimation of viral genomes in females showed that most of them contained 103 RNA copies. Only a few ones appeared to replicate the virus, which reached a viral load of 107 RNA copies (Fig. 3B).
Quantification of Rift Valley fever virus RNA copies in (□) dead and (♦) live females at different days after an infectious blood meal with Clone 13 (
Discussion
RVFV is of increasing concern because of its high potential to cause outbreaks among livestock and humans. RVFV strongly suppresses induction of the antiviral IFN response by means of the NSs protein (Bouloy et al. 2001, Billecocq et al. 2004). The natural isolate Clone 13 expresses a nonfunctional truncated NSs gene and is therefore considered as a potential candidate for a live vaccine. One concern would be the susceptibility of mosquitoes to Clone 13 or more accurately, if Clone 13 induces a transient viremia in vaccinated animals, is a mosquito able to replicate Clone 13 after an infectious blood meal? Moreover, is this vector disturbed by viral infection? Our results showed that Ae. vexans better replicated the two RVFV strains (the avirulent Clone 13 and the virulent ZH548) than Cx. p. quinquefasciatus. Whichever mosquito species was examined, infection with Clone 13 did not induce any important changes in mosquito's life-history traits (survival and reproduction) compared to noninfected females.
Clone 13 was found to be avirulent in rodents and defective in the synthesis of NSs, the nonstructural protein encoded by the S RNA segment (Muller et al. 1995). A virulent strain is able to express a functional NSs protein associated with the nucleus and forming filaments (Swanepoel and Blackburn 1977). The NSs protein interacts with different cellular partners (p44 and SAP30), leading to block IFN-β gene expression at the transcriptional level (Billecocq et al. 2004, Le May et al. 2008) and to suppress host cell RNA synthesis via a decrease in the TFIIH transcription factor concentration in mammalian cells (Le May et al. 2004). If Clone 13 succeeds to induce a viremia in inoculated animals, a disseminated infection would be developed in mosquitoes after feeding and thus be potentially able to transmit virus by bite to a subsequent host. However, even if Ae. vexans and Culex p. quinquefasciatus were both susceptible to Clone 13 (Moutailler et al. 2008), it seems to be unlikely that a mosquito would become infected in nature by feeding on an immunized animal developing a low viremia (e.g., 103 PFU/mL in hamsters during the first 24 h of infection) (Muller et al. 1995). Nevertheless, Egyptian Cx. p. pipiens showed low transmission rates in laboratory, while it was implicated as the principal vector during the 1977–1978 RVF outbreak in Egypt (Meegan et al. 1980). The ecology of Cx. p. quinquefasciatus present in domestic environments colonizing waste water in both artificial and natural containers favors its interaction with domestic animals and humans. Its poor ability to replicate Clone 13 allows limited dissemination of Clone 13. Besides, populations of the floodwater mosquito Ae. vexans are massively produce after a flooding rain. Thus their densities are dependent on rainfalls, which restrict their interactions with animals during a limited period of the year and, consequently, the dissemination of Clone 13.
Even though no cytopathic effects were observed in the vector, reduced fecundity and survival of RVFV-infected Cx. p. pipiens has been demonstrated, related to the extensive tissue and organ infection (Turell et al. 1985, Faran et al. 1987). Females infected with Clone 13 did not show any clearcut changes in mosquito's life-history traits. Indeed, the quantity of resources available for each organism is, in most of cases, limited. So, they have to face trade-offs when they allocate resources to different functions. Consequently, we expected to detect an effect of viral infection on the mosquito's life-history traits, leading the mosquito to adjust its allocation of resources. However, our results showed that infection with Clone 13 did not cause any significant changes in survival and reproduction.
Both mosquito species were able to replicate Clone 13. Nevertheless, a vaccine made using Clone 13 would be less likely to disseminate by vectors. Moreover, attention should be given to the segmented nature of RVFV. Indeed, when a female is infected, the infection is persistent. Thus, if a female contracts an infection through an infectious blood meal containing Clone 13, coinfections could occur with subsequent blood meals. Reassortants would be formed between Clone 13 and virulent strains leading to revert to virulence. Chimeric vaccine should be a promising issue.
Footnotes
Acknowledgments
The authors are grateful to D. Coudrier and C. Tamietti for their technical assistance. We thank F. Rougeon for fruitful discussions. Thanks to two anonymous reviewers for their valuable comments on this paper. We are indebted to F. Lacoste, P. Rassat and T. Fooks for their constant support.
This project was supported by a fellowship to S.M. from the “Fondation de France: Fonds Inckermann,” by the Institut Pasteur and the Arbo-Zoonet project.
Author Contributions
S.M. performed all experiments as part of her Ph.D. work. G.K. participated in some technical aspects. Y.M. contributed to statistical analysis. M.B. supervised the virological aspects of the study. A.B.F. designed the project and wrote the article.
Disclosure Statement
No competing financial interests exist.