
Abstract
The East African Mopeia virus (MOPV) is an arenavirus closely related to the highly pathogenic West African Lassa virus, even sharing the same reservoir rodent host Mastomys natalensis. Because MOPV is not known to cause human disease, it offers a unique alternative for studying Lassa virus transmission. We investigated how habitat, population density, and host characteristics are related to MOPV occurrence in M. natalensis populations in Morogoro, Tanzania. In 3 contrasting habitats, 511 M. natalensis individuals were trapped, 12.1% (58/480 tested individuals) of which tested seropositive for antibodies and 8.4% (41/489 tested individuals) for MOPV-RNA. Although population densities differ among habitats, density and habitat were not significantly correlated to MOPV-RNA or antibody presence. Antibody presence was not significantly correlated with any host characteristics. In contrast, MOPV-RNA presence was inversely related to weight, age, sexual maturity, and body mass index. The model with body mass index as predictor was the best at predicting infection probability. Thirty-five individuals were exclusively MOPV-RNA positive, 52 were exclusively antibody positive, and 6 were both MOPV-RNA and antibody positive. Interpreting these data using experimental infection results from studies on other arenaviruses, this would mean that these infections were very recent, old, and roughly 1–3 weeks after infection, respectively. The higher RNA prevalence in juveniles implies vertical transmission, or that horizontal transmission occurs mainly in this age group due to lack of immunity, higher susceptibility, and/or higher juvenile contact rates. This study demonstrates the strength of combining information on antibody and RNA presence with host characteristics, and how this information can provide valuable insights into transmission dynamics.
Introduction
Unfortunately, because of its biosafety level 4 status, LASV is difficult and expensive to study in the field. The closely related Mopeia virus (MOPV) offers a convenient alternative because it has not been shown to be harmful to humans, and because it occurs in the same reservoir host, the multimammate mouse Mastomys natalensis, be it in East Africa instead of West Africa (Wulff et al. 1977).
In Morogoro, Tanzania, M. natalensis is a common generalist rodent. M. natalensis populations display large demographic fluctuations, both seasonally and interannually, depending on preceding rainfall and population densities. In general, abundance starts to increase in September, corresponding to the end of the dry season and the beginning of the second rainy season, reaching a peak in November. Numbers increase due to a flush of juveniles. This period marks the end of the reproductive season (Leirs 1994, Leirs et al. 1997). The environment may favor the maintenance of the virus directly (humidity, temperature, and UV), or indirectly through the host.
Transmission modes of MOPV are unknown. High-density host populations favor horizontal transmission via individual contacts in the case of density-dependent transmission, often assumed for rodent-borne microparasites (Begon et al. 1999). Another possibility would be vertical transmission, which can, for instance, be seen for Lymphocytic Choriomeningitis virus (Traub 1939) and Junin virus (Vitullo and Merino 1988), in which case high virus prevalence is expected in the youngest animals.
The goal of our study was to investigate how habitat, population density, and host characteristics relate to MOPV occurrence in M. natalensis populations in Morogoro. We sampled individuals in three physically contrasting biotopes across which host density is expected to vary: cultivated gardens, fallow lands, and woodlands. We sought to maximize possible contrasts between biotopes: cultivated gardens may represent a more stable environment in which constant food availability maintains higher population densities than fallow lands or woodlands. Woodlands, with a sandy loose underground and low food availability, are expected to harbor less mice. Gardens may thus exhibit increased maintenance and transmission of the MOPV.
We also expect variations in viral presence related to age. High prevalence in juveniles is expected in case of vertical transmission or due to a lack of immunity. As exposure probability is cumulative over time we expect a positive correlation between antibody (Ab) presence and age.
Materials and Methods
Study sites
Three contrasting biotopes around Morogoro (Tanzania, 6.84°S 37.65°E) were chosen for trapping: (1) young woodland (W), consisting of high (2–10 m) acacia trees, bushy grass undergrowth, and sandy dry soils; (2) fallow fields (F), which are bare clay/loam fields with sparse weed growth, months to a couple of years after crop growth (maize/sunflower); (3) vegetable gardens (G), irrigated year-round to produce vegetables and fruits. We sampled five fallow land sites and five vegetable gardens. Because of very low sample sizes, we sampled six instead of five woodland sites. For the first fallow field, a 1.96 ha trapping grid was set, but because this grid size proved too time consuming, other sites were 1 ha.
Rodent trapping and sampling
Between 13 October and 3 December 2008, two-night capture-mark-removal trapping in 1 ha grids of 100 traps was performed using Sherman live traps baited with a mix of peanut butter and maize scraps. Captured animals were identified morphometrically. Blood samples taken from the retro-orbital sinus were preserved on prepunched filter papers (±15 μL/punch; Serobuvard, LDA 22, Zoopole, France). Weight and body length measurements were taken. Body mass index (BMI; weight/length2) was used as a measure of body condition. Dehydrated eye lens weight was used as a measure of age, calculated using the standard curve constructed by Leirs et al. (1990): a = e (10.46088 + w)/4.35076 where a is age in days and w is weight in mg. Gender and sexual maturity were recorded following Leirs (1994).
Virus and Ab detection
Blood samples were analyzed for both anti-MOPV Abs and MOPV viral RNA (vRNA). Dried blood on filter paper was dissolved in 300 μL phosphate-buffered saline and tested for arenavirus IgG Abs by indirect immunofluorescence assay, using antigens presented by MOPV-infected Vero cells. Viral RNA detection was performed as described in Goüy de Bellocq et al. (2010), using one-step reverse transcription-polymerase chain reaction targeting 320 base pairs of the RNA-dependent RNA polymerase gene (L segment) (Vieth et al. 2007). The MOPV strain in Morogoro is also referred to as Morogoro Virus (Günther et al. 2009).
Statistical analysis
Statistical analyses were performed using “R” (v2.9.0 R Development Core Team 2009) and “WinBUGS” (Lunn et al. 2000).
Confidence intervals for site prevalences (percentage of Ab-positive or vRNA-positive individuals) were constructed in Quantitative Parasitology using Sterne's Exact method (v3.0 Rózsa et al. 2000, Reiczigel 2003).
We wished to capture information regarding the relative abundance of M. natalensis in sampling localities. The study design based on two-night capture allows calculation of the commonly used Lincoln-Petersen (LP) estimator of absolute abundance (Chapman 1951). However, numbers of recaptures were low, in which case sampling noise in recaptures translates to extreme noise in the LP estimator (because of division by small numbers), hence giving very large estimate variances. As absolute abundance estimates are unnecessary when relative abundance is the focus of inference, we explored the efficacy of mean capture probability per hectare, mu, as a proxy for abundance per hectare. Regression shows that mu point estimates capture 92% of the information in LP point estimates, while being much less subject to sampling noise. In addition, the regression outliers correspond to localities where the LP requirement of equal capture probability over visits was very unlikely to be met (due to differing conditions like high rainfall), suggesting that LP information not captured by mu represents noise rather than signal. The mu abundance proxy can be modeled as the rate of a Poisson process, allowing explicit likelihood-based comparison of relative abundances.
Only 100 of 196 traps in site F1 were considered valid for density calculation. To control for edge effects valid traps were chosen to match in numbers the corner and edge classes of traps in each peripheral layer of a 10 × 10 grid. Within these constraints choices were randomized, and 106 valid arrangements were used to estimate edge-effect-controlled capture equivalents for a 10 × 10 grid.
Density differences between sites and biotopes were tested using the G test.
Bayesian generalized linear modeling (GLM) with logit-link function and Bernoulli response distribution was used to analyze infection probability related to density, biotope, gender, sexual maturity, weight, age, and BMI, using Gibbs sampling with 80,000 iterations after 20,000 burn-in iterations, with convergence checked using two chains [see Eddy (2004)]. Because the four latter variables were strongly correlated, we opted for the use of a separate model for each variable, and the comparison of Deviance Information Criterion (DIC) values for assessment of the best predictive variable; the smaller the DIC value, the better the model can predict a replicate dataset with similar structure (Spiegelhalter et al. 2002). Estimated abundances and corresponding shape parameters were used as model priors. Other estimates were given bounded flat priors.
Spatial autocorrelation of site prevalence was analyzed in ArcMap (ESRI 2008) using Moran's I index, which calculates a measure of spatial autocorrelation based on feature locations and values. The index can range from −1 (dispersed) to 1 (clustered) (Moran 1950).
Estimated predictor effects and differences were considered significant when the two-tailed probability (p) was smaller than the commonly accepted 5%.
Results
During 3192 trap nights (# traps × # nights) 555 individual animals were captured, 511 of which were M. natalensis. Analyses and results include data on M. natalensis only. Remaining captured species are described in Goüy de Bellocq et al. (2010), and although some tested positive for anti-MOPV-Abs, they were shown to carry different arenavirus species. Woodland sites W1 and W2 could only be sampled for one night and were therefore not used for density analyses.
Density
The density proxy mu differs significantly between all pairs of ecotopes (G tests for W-G, G-F, and W-F, respectively; df = 1, ΔLL: 150.16, 18.14, 50.67; p = 0, 10−9, 0). There is significant heterogeneity at the level of individual sampling localities (G test for heterogeneity; df = 13; p = 0). Regression of LP on mu indicates that absolute densities are roughly 2/ha in woodland, 46/ha in fallow land, and 118/ha in garden.
MOPV prevalence
Out of 511 trapped animals, 477 were tested for both Abs and vRNA (Table 1).
The first column shows the number of Mastomys natalensis individuals tested with both immunofluorescence assay and RT-polymerase chain reaction. The second and third columns show Ab-negative and Ab-positive samples, respectively.
Ab, antibody; MOPV, Mopeia virus.
MOPV anti-NP Abs and vRNA were present in 12.1% (58/480 individuals tested for Ab presence) and 8.4% (41/489 individuals tested for vRNA presence) of sampled M. natalensis individuals. Six out of 477 (1.3%) tested individuals were positive for both Ab and vRNA. Ab- and vRNA-positive individuals were found in all three biotopes (Fig. 1).
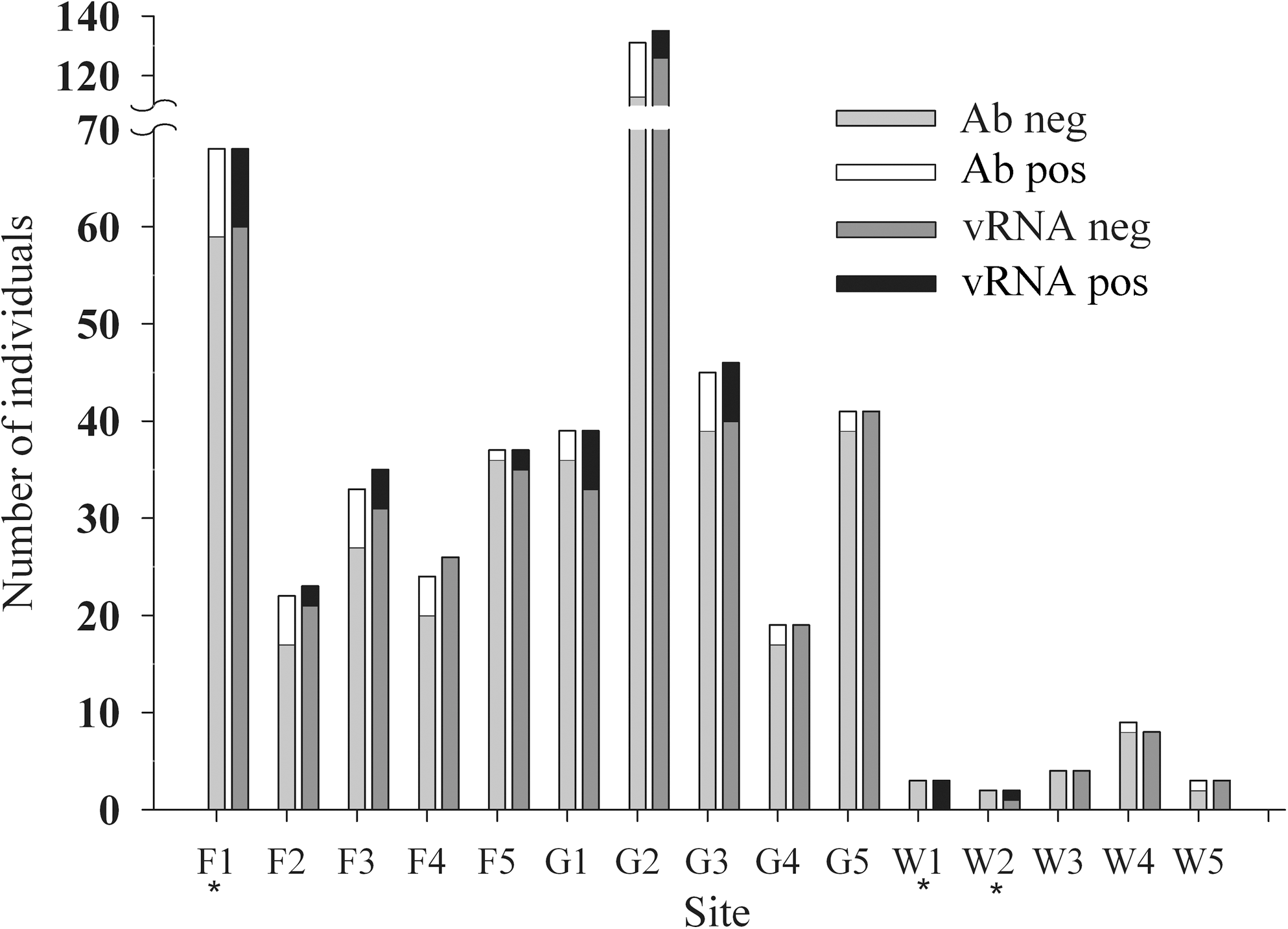
Ab and vRNA prevalence for each sampling site depicted as numbers of individuals. Sites marked with * denote deviances from trapping effort as described in the Materials and Methods section. Ab, anti-MOPV-antibodies; vRNA, MOPV RNA; MOPV, Mopeia virus.
Results from the GLMs are shown in Table 2. There was no significant correlation between biotope and prevalence (Table 2), so data were pooled (i.e., not categorized into three biotopes) to increase power. Because there was no significant spatial autocorrelation of Ab or vRNA prevalence [Moran's I index = 0.19 (Ab), −0.01 (vRNA); Z = 1.58 (Ab), 0.44 (vRNA); p = 0.11 (Ab), 0.66 (vRNA)], the spatial factor was not taken into account in the analyses. For Ab, no significant correlation between infection probability and any of the predictors (density, age, etc.) could be seen, whereas for vRNA there was a significant negative effect of BMI, age, weight, and sexual maturity (Fig. 2).
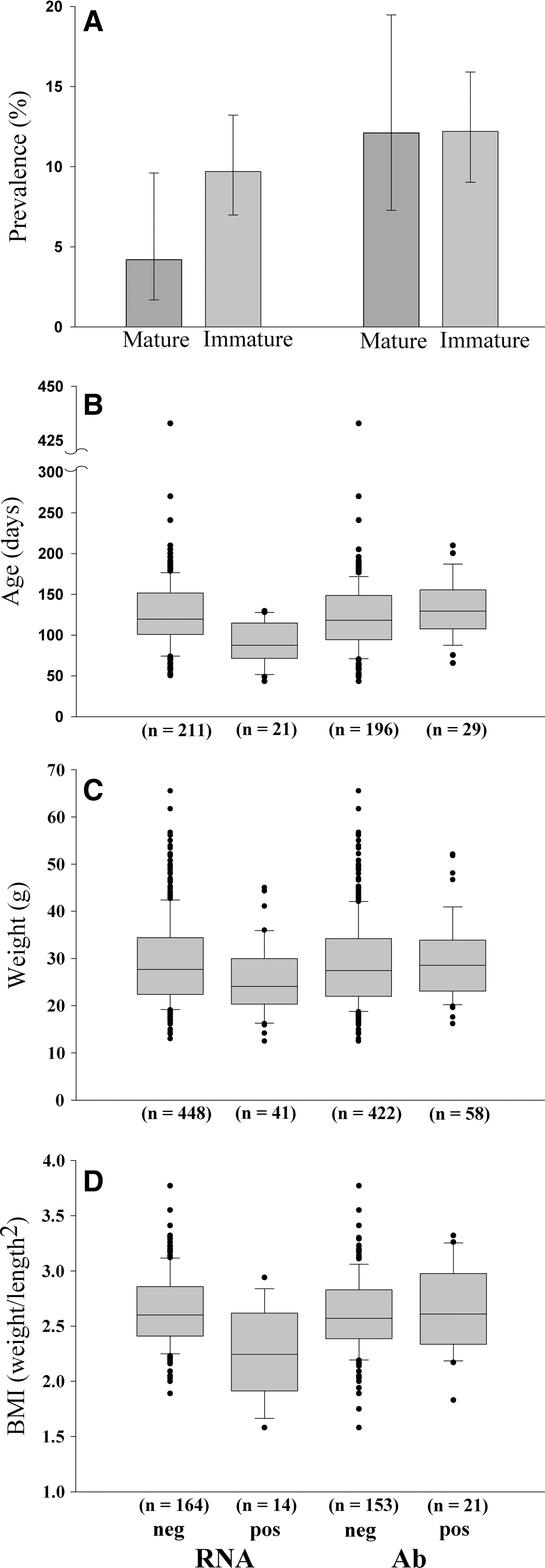
Ab and vRNA prevalence related to host characteristics.
Estimate effects and corresponding standard deviations and confidence intervals are shown in logit scale to allow linear comparison. Effect estimates are considered statistically significant when the 95% confidence interval does not include 0, indicated with the asterisk symbol. DIC values of each model are also given, with a lower DIC value indicating that the model is better at predicting a replicate dataset of a similar structure. Pregnant females were not included in weight and BMI analyses.
DIC, Deviance Information Criterion; SD, standard deviation; CL, confidence limit; n, sample size; PC, principal component; BMI, body mass index.
The model with BMI as predicting variable had the lowest DIC for both vRNA and Ab. Because BMI was strongly correlated with age (t 177 = 9.13, R = 0.56, p < 0.0001), including both in a regression model would lead to multicolinearity problems. To investigate effects of both BMI and age, we first performed a principal component (PC) analysis of BMI and age. This resulted in two components where PC1 had loadings 0.707 for BMI and 0.707 for age, and PC2 had −0.707 for BMI and 0.707 for age. The first principal component thus reflects a weighted average of age and BMI, whereas the second is the contrast between both. Higher values of PC1 indicate on average higher BMI and age, whereas higher values of PC2 indicate low BMI relative to the observed age. PC1 accounted for 78% of variance, and PC2 for 22%. These two principal components were then used in a Bayesian regression model relating prevalence to PC1 and PC2, where PC2 could now be largely interpreted as the effect of BMI independent of age. For vRNA, the effect of PC1 was significant, whereas the effect of PC2 was not (Table 2). For Ab, none of the principal components showed a significant effect on prevalence.
Figure 3 shows the age composition of all tested individuals. Although age was not significantly correlated to Ab occurrence, Ab prevalence shows a positive trend within age groups when categorized into months (GLM, logit link, age effect = 0.24, df = 6, p = 0.107). Supporting the GLM results, vRNA-positive individuals are mainly found in young individuals, reaching a peak at 3 months. After 3 months, the proportion of vRNA-positive individuals decreases (GLM, logit link, age effect = −0.82, df = 6, p < 0.001), with no vRNA-positives older than 5 months. Thirty-five animals (7.3%) were Ab-negative and vRNA-positive (Ab−/vRNA+), ranging in age from 43 to 130 days (N tested = 17/35). Six animals (1.3%) were Ab+/vRNA+, aged 88 to 115 days (N tested = 4/6), and 52 (10.8%) were Ab+/vRNA−, aged 66 to 210 days (N tested = 25/52).
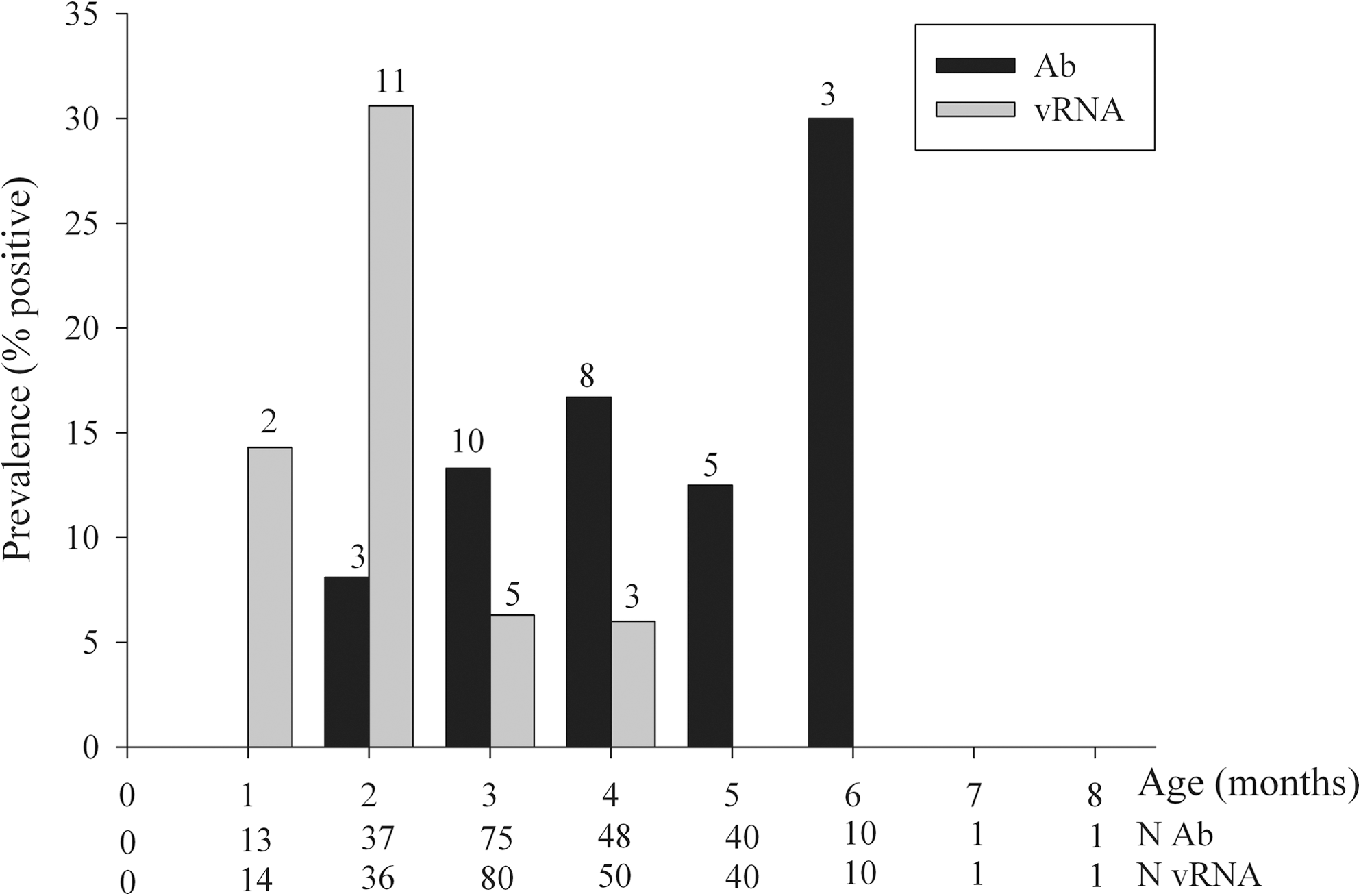
Age related to Ab and vRNA presence. Age is given in months, with 0–30-day-old individuals in the 1-month-old age category, 31–60-day-olds in the 2-month category, and so on.
Discussion
In the present study, we tested how environment, host density, and host characteristics affect the occurrence of MOPV or anti-MOPV-Abs. We found no significant relationship between Ab presence and any measured variable. Viral RNA was not significantly correlated with gender, density, or biotope, but did correlate inversely with all variables related to age.
Abiotic environment
There has been very little research on the abiotic factors determining arenavirus occurrence and survival outside the host (Sinclair et al. 2008). It is known that biological half-lives of aerosolized LASV at both 24°C and 32°C ranges from only 10 to 54 min (Stephenson et al. 1984). On a larger scale, the occurrence of LASV in West Africa has been linked to rainfall regime and quantity, independent of host occurrence (Fichet-Calvet and Rogers 2009).
This study revealed no difference in prevalence between the three abiotically contrasting biotopes, and this suggests that the small-scale abiotic environment of these biotopes is not an important direct determinant of virus occurrence, even if on a larger spatial scale there is such a relation. The focus should therefore lie on the role of the host in determining virus presence.
Body mass index
One of the results that stand out strongly is the significant role of body condition in predicting the presence of MOPV or Abs. BMI can be related to age as well as to body health, in that young animals generally have a lower BMI than old animals (also shown for voles in Olsson et al. 2002), and that unfit animals have a lower BMI (Willett 1998). Because we also found a significant negative correlation between vRNA prevalence and age, weight, and sexual maturity (all correlated with age), the significance of BMI is probably for a large part due to age effects. Even though the effect of PC2 (the effect of BMI largely—but not entirely—independent of age) was not significant, the fact that the model with BMI was a better predictor for MOPV prevalence than the other models still suggests body condition effects unrelated to age. This same pattern has also been found previously by Demartini et al. (1975).
Age
The proportion of Ab-positive individuals increases with age, whereas the proportion of vRNA-positive individuals decreases (Fig. 3). This pattern is exactly what would be expected if probability of infection, followed by Ab production and virus clearance, is cumulative over time, or if infection induces higher mortality. Increasing Ab prevalence with age has been seen for other arenaviruses (Demby et al. 2001, Mills et al. 1994, 2007), implying that horizontal transmission followed by Ab production may be common among arenaviruses.
LASV RNA has been detected in very young M. natalensis, including 8–10 days olds, and prevalence has been seen to increase with age until 4 months (Fichet-Calvet et al. 2008), which strongly suggests vertical transmission combined with horizontal transmission among juveniles. Although the youngest animal captured during our study was 43 days, the similar pattern of vRNA prevalence in our study indicates a similar mode of vertical and horizontal transmission, a hypothesis further supported by the low Ab prevalence in juveniles that suggests an absence of maternal Abs.
The high vRNA prevalence in juveniles fits the over-representation and increased dispersal activity of juveniles during the annual period of high population density (Leirs 1994). It could be related to these higher densities, yet no significant correlation between site density and MOPV prevalence was found. It thus appears to be the case that juveniles are more susceptible, be it through lack of immunity, higher intrinsic physiologic susceptibility, and/or higher contact rates than among older animals.
Even in woodland, with its very low densities, high Ab and vRNA prevalences were found, which hints at deviation from density-dependent transmission. Importantly (especially when extrapolated to LASV), this finding warns that there is an infection risk to humans in all biotopes where M. natalensis occurs, which is lower yet still present in low-density biotopes.
Transmission
To interpret our serological data in an ecological context and relate it to transmission dynamics, one would ideally use results from experimental infection studies performed with MOPV on M. natalensis. Unfortunately, no such studies are available. There does exist a study by Walker et al. (1975) on LASV in Mastomys coucha (and not the natural reservoir host although it was, mistakenly (Kruppa et al. 1990), considered to be M. natalensis in the study), which found viraemia from 7 to at least 74 days postinfection (PI) in neonatally infected animals and on day 12 PI for animals infected as adults. Abs were detectable from day 50 PI but often never (Walker et al. 1975). Because of the low sample size and the use of a related but nonreservoir host, these results should, however, be interpreted carefully. To our knowledge there is one other experimental arenavirus infection study by Fulhorst et al. (2001), ?tlsb=0w> on the New World Whitewater Arroyo Virus. Here, virus in blood was detectable from 15 to 55 days and 7 to 21 days PI in individuals infected on day 1 and 120, respectively, and Abs from 21 to at least 164 days PI and 7 to at least 56 days PI in animals infected on day 1 and 120, respectively. Although virus can usually not be detected in blood for a long time, it is often detectable in organs, urine, and saliva for longer periods (especially in early infections) during which viral particles can probably still be shed (Walker et al. 1975, Demby et al. 2001, Fulhorst et al. 2001). Additionally, several experimental studies have independently shown that arenavirus infection of newborn and young rodents can lead to a lifelong chronic infection, whereas it is usually transient when infected at an older age (Walker et al. 1975, Johnson 1981, Vitullo et al. 1987, Fulhorst et al. 2001).
Using all this information to interpret our results, this would mean that the 6 (1.3%) Ab+/vRNA+ individuals in our study have occurred roughly a few weeks before sampling, whereas the 35 (7.3%) Ab−/vRNA+ individuals are even more recent infections. The 52 (10.8%) Ab+/vRNA− individuals should then be interpreted as older infections where the virus has been cleared, at least from the blood.
Linking the interpreted results to age and changes in population structure now provides valuable insights about transmission dynamics. All animals with recent infections (Ab−/vRNA+ and Ab+/vRNA) in our study were 1.5 to 4 months old, and this is suggestive of vertical transmission (although tentatively, as no animals younger than 43 days were captured) and/or horizontal transmission rates that are higher among juveniles, and in any case indicates that this age group, at this time of year, is the most important for virus transmission. If it is correct that young animals indeed more readily develop chronic infections, then this age group would have an additional important role for virus persistence. Further, because the demographic structure of M. natalensis populations changes throughout the year, it can be expected that transmission rates and infection probability toward humans will vary accordingly, with peaks expected to coincide with periods of high juvenile abundance.
Footnotes
Acknowledgments
The authors thank two anonymous reviewers for helpful comments that improved the article. Fieldwork was performed with the help of the enthousiastic staff at SUA PMC, in particular Khalid Kibwana and Shabani Lutea. Thanks to Stefan Van Dongen and Stuart JE Baird for statistical help, and to Katrien Tersago for useful comments. This study was funded by the University of Antwerp (JGB grant Nr. 2964) and the Fund for Scientific Research-Flanders (FWO) (JGB grant Nr. 1.5.180.09). JGdB is a postdoctoral fellow with FWO. Travel expenses of BB were partly financed by VLIR.
Disclosure Statement
Authors declare that no competing financial interests exist.