
Abstract
Antimicrobial peptides are an essential component of the insect immune system. One of the most ubiquitous is defensin, which has been identified in all examined insect orders. Triatoma brasiliensis (Heteroptera, Triatominae), the main Trypanosoma cruzi vector in semi-arid regions of north-eastern Brazil, expresses def1, a defensin encoding gene, predominantly in the anterior region (cardia and stomach) of the midgut. In the present study, we compared the transcript abundance of T. brasiliensis def1 in the anterior (stomach) and posterior midgut (small intestine) regions of naïve bugs with those infected with a familiar T. cruzi isolate. In the stomach, only slight differences between the two insect groups were visible, whereas in the small intestine wide differences (up to 9.6-fold) between infected and noninfected bugs become apparent. The highly increased def1 transcript abundance at 20 days after the infective blood meal might be a response to the T. cruzi infection, suggesting a potential function of intestinal defensin in the parasite population control.
Defensins are cationic antimicrobial peptides occurring in many insect groups and important effector molecules of the insect innate immune system (Boulanger et al. 2006). The first triatomine defensins and respective genes have been described in Rhodnius prolixus and named defensin A, B, and C (Lopez et al. 2003). Using either Northern blot analysis or real-time quantitative polymerase chain reaction (PCR), a strong defensin gene upregulation has been shown in the intestine and fat body, after inoculation of bacteria into the hemocoel.
In the present study, T. brasiliensis fifth instar larvae were artificially fed with blood containing T. cruzi, which was previously isolated from naturally infected T. brasiliensis. Def1 encoding mRNA levels—encoding an intestinal defensin—were analyzed by semi-quantitative reverse transcription (RT)-PCR in uninfected and T. cruzi infected insects at different feeding conditions.
T. cruzi strain TBRA/BR/1999/JCA3 (TcII) obtained from naturally infected peridomiciliar T. brasiliensis, in Piauí, north-east Brazil, had low passages in medium culture and was stored in liquid nitrogen, until use (Herrera et al. 2003). Fifth instar larvae of T. brasiliensis, kindly provided by Prof. J. Jurberg (Fiocruz, Rio de Janeiro), were artificially fed with sterile noninactivated citrated rabbit blood and blood containing 1 × 106 cells/mL through latex membranes (Garcia et al. 1989).
For analysis of T. brasiliensis def1 transcript abundance, stomach and small intestine tissues of bugs (n = 10) at 5, 10, and 20 days after infection were dissected. During the dissections, the total number of parasites in the intestine was determined using Neubauer hemocytometer. Total RNA was isolated and measured as previously described (Araújo et al. 2006). Semi-quantitative RT-PCR and cloning were carried out as previously described using the specific oligonucleotide primers RT-Def1-F (5′-TCCCTTCTACCCAGCAAATATTATG-3′) and RT-Def1-R (5′-GTCTTATATCTTCTTCTTTAGCAATTGG-3′), designed according to T. brasiliensis def1 [GenBank:AY641574] (Araújo et al. 2006, Waniek et al. 2009).
In the stomach lumen, no parasites were observed. The number of parasites colonizing the small intestine increased over time. At 5 days after feeding, the small intestine was colonized by a mean of 3 × 102 parasites/insect. The parasites concentration increased on average to 1 × 104 parasites/insect at 10 daf and reached its maximum of 4.6 × 104 parasites/insect at 20 daf.
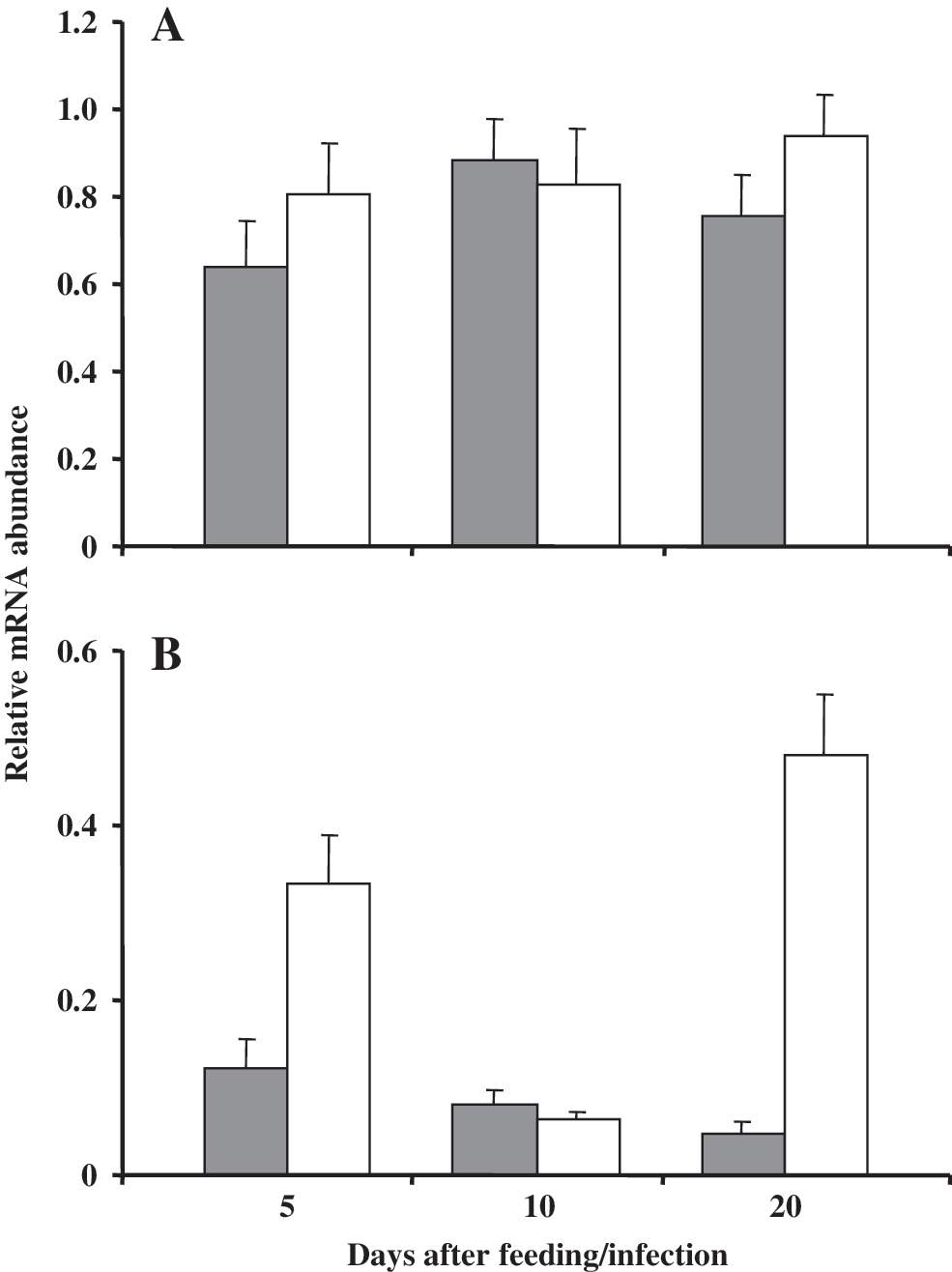
To verify the primer specificity, 13 clones were sequenced. All of them corresponded in their nucleotide sequence with that of T. brasiliensis def1-cDNA, confirming stringent primer specificity. All amplification products resulted in a single band, which confirmed complete gDNA digestion. In negative controls lacking cDNA, carried out for each sample, no amplification products became visible. The def1-mRNA concentrations were always significantly higher in the stomach, the region of highest symbiont concentration, in comparison to the small intestine (Fig. 1). In the stomach of infected and noninfected insects, def1 was constitutively abundant. The means of def1-mRNA levels slightly but not significantly increased in T. cruzi infected bugs at 5 and 20 days after feeding and not in infected bugs 10 daf, respectively (Fig. 1A). In comparison with the stomach, the small intestine tissue concentrations of def1 transcripts were clearly more variable (Fig. 1B). In the small intestine of uninfected bugs, the def1 transcript abundance reached its peak at 5 days after feeding decreasing 10 and 20 daf, whereas in infected bugs the def1-mRNA concentration strongly fluctuated. At 5 days after feeding, def1-mRNA was significantly higher in infected bugs (2.8-fold, p < 0.005). At 10 days after feeding, def1 concentration was almost identical in both groups. At 20 days after feeding, the clearest difference between the two insect groups occurred, with a 9.6-fold def1-mRNA increase in T. cruzi infected insects and highest significance level (p < 0.00001). At this time, the relative def1-mRNA abundance of infected insects in the small intestine was comparable to that detected in the stomach (Fig. 1).
Semi-quantitative reverse transcription polymerase chain reaction analysis of Triatoma brasiliensis def1 with total RNA from the stomach (
In the stomach, T. cruzi can be found only a short time after infection due to its confrontation with hostile components (e.g., hemolytic factors, lectins) (Kollien and Schaub 2000, Garcia et al. 2007). In this midgut region, def1-mRNA abundance remains comparatively constant, suggesting a probable function of mature Def1 in the control of luminal symbiotic bacteria (Araújo et al. 2006). However, the number of symbionts is significantly reduced only in the small intestine, suggesting that additional factors such as low pH or high concentrations of proteolytic enzymes—only present in the posterior midgut region—are crucial (Kollien and Schaub 2000, Eichler and Schaub 2002). The def1-mRNA levels in the small intestine of uninfected bugs (Fig. 1B) are similar to the level previously described, much lower than in the stomach at 5 days after feeding, slightly decreasing in the following days (Araújo et al. 2006). On the other hand, in the small intestine of T. cruzi infected bugs, the def1-mRNA levels fluctuated strongly. It is possible that the different growth behavior of intestinal symbionts and T. cruzi (peaks at 5 and 20 daf, respectively) influences the def1-mRNA levels (Eichler and Schaub 2002). Highest numbers of T. cruzi in the small intestine at 20 days after feeding might modulate the def1 abundance in this midgut region. Therefore, high def1 abundance in the stomach might reflect its adaptation to synthesize high quantities of defensin due to symbiont multiplication, whereas the small intestine, in which intestinal bacteria are less abundant, seems to be more sensitive to def1 expression induction by organisms occasionally inhabiting the intestine, leading to a broader def1 transcript abundance range.
Once infected, triatomines show high parasite densities in the intestine, which are only reduced after long periods of starvation (Kollien and Schaub 2000, Araújo et al. 2007). Thus, the elevated def1-mRNA abundance in the small intestine might be a response to T. cruzi infection in this midgut region, in which activated Def1 might act as a signaling molecule in the immune cascade (Boulanger et al. 2006). If mature Def1 is active against trypanosomes, its function might be restricted to control the parasite population and not be responsible for its elimination. However, a selection of the fittest T. cruzi strains by intestinal defensin cannot be excluded. Up to what extent the parasite population is directly affected by higher defensin levels, or whether the increase of def1-mRNA is a sign of the T. cruzi infection, can only be verified in further defensin activity or RNA interference tests. Most probably, there are several further immune factors that influence T. cruzi population in the intestine of Triatominae.
Footnotes
Acknowledgments
To Prof. O. Fernandes, Prof. D. Valle, and Prof. V. Bongertz (Fiocruz, Rio de Janeiro) for technical assistance and English revision of the manuscript, respectively. The present work was financially supported by Fundação de Amparo á Pesquisa do Estado do Rio de Janeiro (FAPERJ/“Cientistas do Nosso Estado”: E-26/100.456/2007), EC contracts 223034 chagas Epi Net and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq/“Edital Universal”: 472276/2006-9 and PDJ: 152069/2006-2). C.A.C.A. is a CNPq Research Fellow (PDJ: 151187/2009-6), and P.J.W. is an FAPERJ Research Fellow (E-26/152.913/2005).
Disclosure Statement
On behalf of the authors, P.J. Waniek certifies that all the authors actively contributed to the study and approved the final version of the article. No competing financial interests exist.