
Abstract
Following veterinary alerts of Rift Valley fever (RVF) in the districts of Fianarantsoa I and II in November 2008 and in the district of Ambalavao in April 2009, entomological and virological investigations were carried out to identify the mosquito species that could act as RVF virus (RVFV) vectors in the region. A total of 12,785 adult mosquitoes belonging to 5 genera and 21 species were collected. After identification, mosquitoes were pooled by species, sex, and female status (fed or unfed) and then stored at −80°C. Of 319 pools of unfed monospecific female mosquito tested by real-time RT–polymerase chain reaction, RVFV was detected in 1 pool of Anopheles coustani, 5 pools of An. squamosus, and 2 pools of Culex antennatus mosquitoes. The virus was isolated in mosquito cell lines from two of the five Real Time-RT–polymerase chain reaction (real time-RT-PCR) positive pools of An. squamosus mosquitoes. From the eight RVFV strains detected, partial S, M, and L genome segments sequences were obtained. The phylogenetic analysis of these sequences showed that the strains circulating in mosquitoes were genetically close to those that circulated in livestock and humans during RVF outbreaks in 2008 and 2009. This study, therefore, provides strong evidence that An. squamosus, An. coustani, and Cx. antennatus could play a role as vectors of the RVFV during the disease outbreaks in 2008–2009. Bioecological, genetic, and RVF transmission studies on these three mosquito species are needed to address this question and thus improve prevention and control of future RVF outbreaks in Madagascar, where these species are present.
Introduction
Infections may also result from bites of infected insects such as mosquitoes. Six mosquito genera (Aedes, Culex, Mansonia, Anopheles, Coquillettidia, and Eretmapodites) including about 40 species were described as naturally infected by RVFV (EFSA 2005, FAO 2008). Members of Culex species play a major role in RVFV transmission in epizootic/epidemic cycles (Meegan 1981, House et al. 1992, Zeller et al. 1997, Diallo et al. 2005) and members of Aedes species, specially those of the subgenus Neomelaniconion (Ae. circumluteolus, Ae. palpalis, Ae. lineatopennis), are involved in the maintenance cycle (McIntosh et al. 1972, 1983, Davies and Highton 1980, House et al. 1992, Zeller et al. 1997) as a vertical transmission is observed (Fontenille 1989).
In Madagascar, the RVFV was first isolated from pools of mosquitoes of the species Anopheles coustani, An. fuscicolor, An. pauliani, An. squamosus, Culex simpsoni, Cx. vansomereni, Cx. antennatus, Cx. quinquefasciatus, Cx. annulioris, Cx. univittatus, Mansonia uniformis, and Coquillettidia grandidieri captured in 1979, during the rainy season in the rain forest of Perinet (Clerc and Coulanges 1981). Nevertheless, the precise implication of each species in the RVFV transmission was not demonstrated as all the pools, except that of Mansonia uniformis, were made up of a mixture of different species.
The first epizooties of RVF were described in 1990 and 1991 during which period several thousand mosquitoes were collected and tested by virus isolation on cell cultures. Nevertheless, no mosquito had been found infected (Morvan et al. 1991a, 1991b, 1992a, 1992b).
In 1993, entomological surveys carried out in parallel to human serological surveys at Antananarivo resulted in the capture of 61,298 mosquitoes with 43% being Aedes circumluteolus. In the homogenates of these mosquitoes, no RVFV-antigen was detected (Morvan et al. 1993). Between 1996 and 1998, 102,637 mosquitoes comprising 61% of Anopheles sp., 34% of Culex sp., 2.4% of Mansonia sp., 2% of Aedes sp., and a low proportion of Aedemomyia sp., Coquillettidia sp., Eretmapodites sp., Mansonia sp., and Uranotaenia sp. were captured. No RVFV was isolated from the 6525 pools tested (Zeller et al. 1998).
The existence of a human sentinel surveillance network set up by the Malagasy Ministry of Health in collaboration with the Institut Pasteur de Madagascar (Randrianasolo et al. 2010) since March 2007 allowed the detection of RVFV in Madagascar in February 2008. This first detection was followed by an alert launched by the Ministry of Health. Overall, during January 28 to June 15, 2008, 476 human suspected cases (of which 19 died) from 15 districts, mostly from the central highlands, were reported. RVFV infection in humans and animals was confirmed (Andriamandimby et al. 2010).
A few months later, during the following rainy season, a veterinary RVF alert from the districts of Fianarantsoa I and II in November 2008 and from the district of Ambalavao in March 2009 was confirmed by the detection of RVFV in samples collected from affected cattle in these areas (Andriamandimby et al. 2010). Subsequent entomological investigations were undertaken in the districts of Fianarantsoa I and II, and Ambalavao in December 2008 and April 2009, respectively. The objective of these investigations was to identify the mosquito species that could act as RVFV vectors in the region. In this article, we report the findings derived from these latest investigations.
Materials and Methods
Study area
Three districts of the Haute Matsiatra region located in the Central Highland of Madagascar (Fianarantsoa I, Fianarantsoa II, and Ambalavao) have been explored (Fig. 1). All collection sites were located at an altitude ranging from 1050 to 1205 m. Fianarantsoa I was the largest agglomeration of the region, whereas the other two districts investigated were located in a rural area. Livestock, including imported dairy cows and common zebus (the predominant animals), was present at all studied sites.
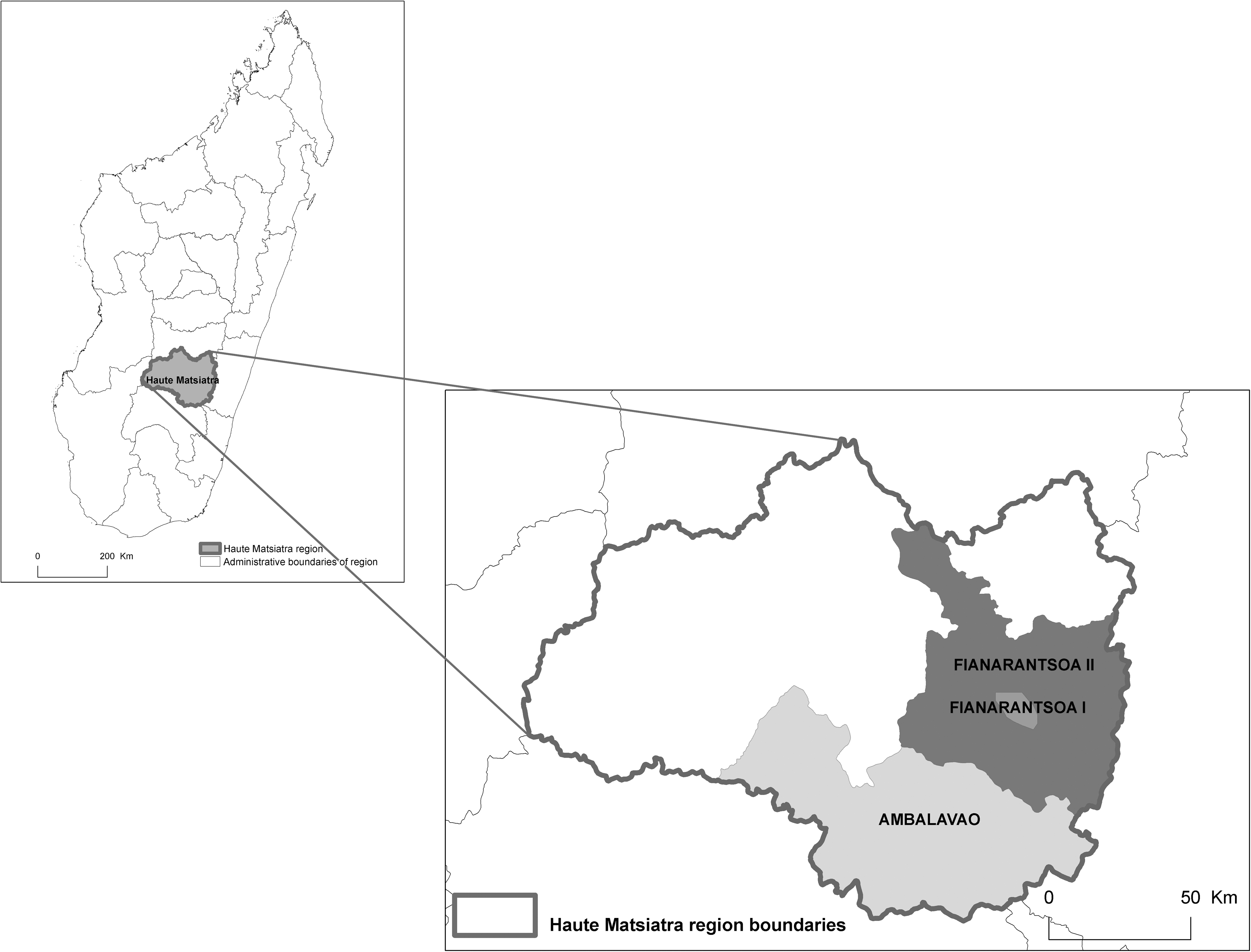
Geographical situation of the three study sites. The three districts are located in the Central Highland of Madagascar at an altitude ranging from 1050 to 1205 m.
Mosquito sampling
Adult mosquitoes were collected in net animal-baited traps, CDC light traps (with or without CO2) set near or next to livestock, and with oral aspirators in mosquito-breeding places. The traps were run for two nights in each study site from 5.30 pm to 5.30 am.
Mosquitoes were either immediately identified in the field or frozen in liquid nitrogen and then later identified morphologically in the laboratory on a chill table. After identification, mosquitoes were sorted according to species, sex, and female status (fed or unfed) and then pooled and stored at −80°C.
Virus detection and molecular characterization
Pools of 1–38 unfed monospecific female mosquitoes were placed in 2-mL microtubes. They were then homogenized in 0.5–1.5 mL cell culture medium (MEM) (containing 40% fetal bovine serum, 2 mM
Viral RNA was extracted from supernatants using TRIzol LS reagent (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions. The molecular detection of the virus was performed using real-time RT–polymerase chain reaction (real time-RT-PCR) as described by Weidmann et al. (2008). The minimum level of detection was 25 copies of transcript RNA per assay.
Virus isolation was performed on mosquito cell lines (AP61) using supernatants from RVFV real-time RT-PCR–positive pools and virus identification was performed by an indirect immunofluorescence assay that used mouse RVFV immune ascitic fluid (Reynes et al. 1994).
To characterize the circulating RVFV strains, parts of the S, M, and L segments were amplified and sequenced as previously described (Andriamandimby et al. 2010).
The amplification products were sequenced on both strands by Cogenics (Meylan, France). Unverified sequences and chromatograms were compared and corrected when needed. Sequences from the same segment were compared when aligned and a phylogenetic analysis was conducted with MEGA version 4 software (Tamura et al. 2007). The partial S, L, and M sequences obtained in this study are available from GenBank under accession numbers GU135840–GU135863.
Results
Mosquito sampling
A total of 12,785 adult mosquitoes, belonging to 5 genera and 21 species, were collected (Table 1). In Fianarantsoa I, among the eight mosquito species collected, Cx. antennatus (90. 45%, n = 1355) followed by An. gambiae s.l. (4.47%, n = 67) was predominant. In Fianarantsoa II, 5745 adult mosquitoes belonging to 4 genera and 18 species were captured. The species An. squamosus (73.32%, n = 4212) and Cx. antennatus (15.82%, n = 909) were most abundant. In Ambalavao, 5542 adult mosquitoes belonging to 4 genera and 16 species were collected, and An. squamosus (74.38%, n = 4122) was the predominant species.
N, number per species; %, percentage per species; RVFV, Rift Valley fever virus.
RVFV detection and isolation
We tested a total of 319 pools, including 59, 126, and 134 pools from Fianarantsoa I, Fianarantsoa II, and Ambalavao, respectively (Table 1). RVFV RNA was detected in eight pools: two pools of Cx. antennatus from Fianarantsoa I, one pool of An. coustani and two pools of An. squamosus from Fianarantsoa II, and three pools of An. squamosus from Ambalavao (Table 1). Further, we inoculated these eight RVFV RT-PCR–positive pools on AP61 cells and isolated RVFV from two An. squamosus pools (FI-79 from Fianarantsoa I and AL-51 from Ambalavao).
RVFV phylogenetic and sequence analyses
Verified partial S sequences encompassing 627 nucleotides (nt positions 49–675 of the coding domain) were obtained from the RVFV strains detected in the eight pools. A phylogenetic analysis was applied using the neighbor-joining (NJ) method with the Kimura 2 parameter model to these 8 sequences and to the corresponding part of (1) representative Malagasy sequences of strains that circulated in Madagascar in 2008–2009, especially those that circulated in the Fianarantsoa districts (Andriamandimby et al. 2010), (2) the 33 complete S sequences described by Bird et al. (2007), and (3) 6 sequences representative of the lineages Kenya-1, −1a, and −2, which were recently described during the 2006–2007 Kenyan outbreak (Bird et al. 2008). Although the bootstrap values did not support an unambiguous phylogenetic classification, the analysis showed that the sequences obtained from the eight Malagasy mosquitoes RVFV strains were identical or close to the sequences obtained from human and cattle RVFV strains that circulated in the Haute Matsiatra region in Madagascar during the outbreaks in 2008 and 2009 and to sequences obtained from Kenyan livestock strains that circulated in 2006–2007, especially to Kenya-1 and −1a lineages (Fig. 2). No amino-acid change was associated with these eight sequences when compared with the partial S sequences of the Kenya-1 and −1a lineages.
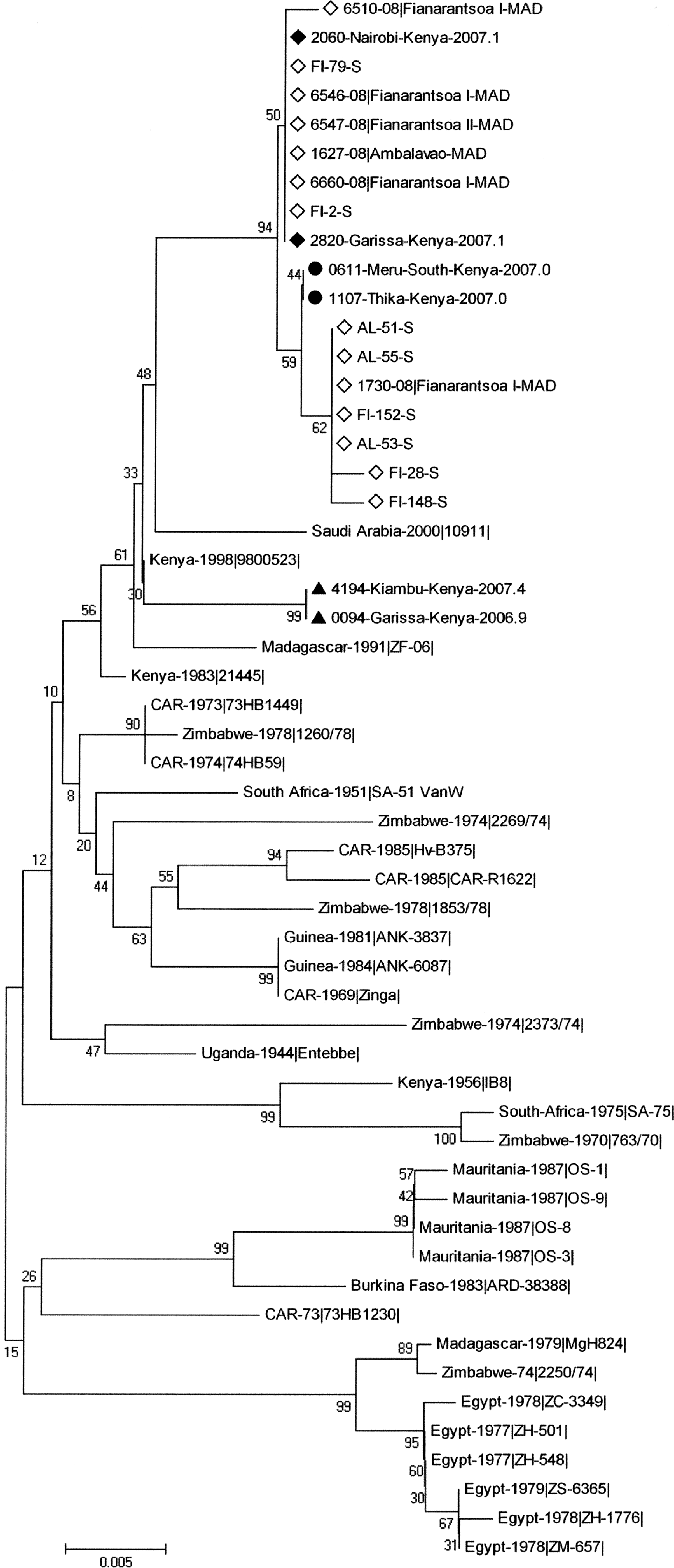
Phylogenetic tree based on the partial S sequences (627 nucleotides) from 33 Rift Valley fever virus (RVFV) strains described by Bird et al. (2007), from six sequences representative of the three Kenyan 2006–2007 RVFV lineages described by Bird et al. (2008), from one Malagasy cattle RVFV strain that circulated in 1991, from six representative Malagasy cattle and human RVFV strains that circulated in 2008–2009 (GenBank accession numbers GQ443166, GQ443168, and GQ443172–GQ443175; Andriamandimby et al. 2010), and from the eight mosquitoes RVFV strains described in this article. (FI 79-S, FI 2-S, AL 51-S, AL 55-S, FI 152-S, AL 53-S, FI 28-S, FI 148-S). Bootstraps percentages (from 1000 resamplings) are indicated at each node. (♦) Sequences from the 2006–2007 Kenyan-1 lineage; (•) sequences from the 2006–2007 Kenyan-1a lineage; (▴) sequences from the 2006–2007 Kenyan-2 lineage; (◊) 2008–2009 Malagasy sequences.
Partial M sequences (nt positions 781–1536 of the coding domain) and partial L sequences (nt positions 1276–1839 of the coding domain) were available for the eight strains. Phylogenetic analysis performed as described above and including the M or L sequences of the strains used for the S analysis confirmed the result obtained with the partial S sequences (data not shown). No amino-acid change was associated with the eight partial L sequences when compared with the batch of L sequences used in the phylogenetic analysis. When compared with the batch of M sequences used in the phylogenetic analysis, six of the eight partial M sequences exhibited one amino-acid change at position 368 of the coding domain (at position 216 in the Gn protein). This change, L368Q, was observed in all the eight strains except the one associated with An. coustani and one of the two associated with An. squamosus.
Discussion
Three species of mosquitoes, An. coustani, An. Squamosus, and Cx. antennatus, were found infected by RVFV in the present study. These three mosquito species were part of the mosquito pools (made up of a mixture of different species) tested and found infected by RVFV in 1979 in Madagascar (Clerc and Coulanges 1981). Our results provide further evidence for the potential strong involvement of these three species in RVFV transmission in Madagascar. Two of the three species have been also found to be infected in other countries affected by RVF: originally, An. coustani was found naturally infected in Zimbabwe in 1969 and Cx. antennatus was found infected with the same virus for the first time in Nigeria in 1967 and then in Kenya from 1981 to 1984 (Linthicum et al. 1985, EFSA 2005). The vector competence of Cx. antennatus was soon demonstrated in laboratory studies (EFSA 2005). The combined evidence indicates that these species are serious RVFV vector candidates in Madagascar.
The RVFV partial sequences we obtained from the mosquitoes were identical or close to the ones we got from cattle and humans in the same areas. This sustains the probable implication of the mosquito species found infected during the Malagasy outbreak in 2009. Six of the eight RVFV strains we detected exhibited one original amino-acid change at position 368 of the coding domain (at position 216 in the Gn protein) when compared with the batch of sequences we used in our analysis. Four distinct immunogen antigenic determinants have been localized along the RVFV Gn glycoprotein (Keegan and Collett 1986). The L238Q (M coding domain position) = L216Q (Gn position) mutation we described is out of these four domains. However, it is close to domain II (amino acids 229–239 of the Gn protein). We do not know how it could interact with this domain. This change in L368Q corresponding to the substitution of a hydrophobic amino-acid by a polar amino-acid may modify the tertiary structure of this glycoprotein and consequently may affect the protective immune response or the recognition abilities of receptors sites on susceptible cells (we do not find any information about them). Further molecular analysis and phenotypic assays (including the study of the susceptibility to the strains of both mosquito species carrying the mutation and species without mutation) are planned.
Studies undertaken in the first half of the 20th century on the geographical distribution of the species of the genus Anopheles in Madagascar showed that An. squamosus and An. coustani were present in all areas of Madagascar (Grjebine 1957). Fontenille et al. confirmed this spatial distribution and showed that Cx. antennatus shares the same habitat with both An. coustani and An. squamosus in 1989. Moreover, during entomological investigations (without virus detection) we conducted at and around the locations of confirmed cases of RVF in the north, the highlands, and the fringe of the highlands of Madagascar in 2008, Cx. antennatus was present in high numbers at all study sites simultaneously with An. squamosus and An. coustani, except in the capital Antananarivo (J. Ratovonjato, unpublished data). Recent serological investigation in humans and livestock suggests that RVFV has circulated in all regions of Madagascar (Andriamandimby et al. 2010, Jeanmaire et al. 2010). Consequently, the wide distribution of these three mosquito species may explain the occurrence of RVF transmission in the entire country.
Among the 14 other mosquito species we identified during our study (Table 1) and those previously described in Madagascar (Grjebine 1957, 1966, Fontenille 1989), five species, Cx pipiens, Cx poicilipes, Cx. tritaeniorhynchus, Cx.univittatus, and Mansonia uniformis, were previously found to be naturally infected by RVFV in Madagascar and/or in other countries (EFSA 2005). These five species were not found infected in our study. However, their role in the transmission of RVFV in Madagascar cannot be ruled out. Questions remain on the implication of mosquito species we did not collect during the RVF outbreaks in Madagascar in 2008–2009, especially species from the genus Aedes and in particular from the subgenera Neomelaniconion and Aedimorphus, previously described in Madagascar (Fontenille 1989, Fontenille and Rodhain 1989). Some are known to be RVFV vectors in Africa, that is, Ae. circumluteolus, Ae. palpalis, and Ae. dalzieli (Fontenille et al. 1998, EFSA 2005). These species could have played a role in the Malagasy RVF outbreak foci, but because we trapped mosquito during the night we might have a bias in our sampling because Aedes species are daytime feeders.
Nevertheless, taken together that (1) the most abundant species we collected at three study sites were An. coustani, An. squamosus, and Cx. antennatus and (2) the close genetic relationships between the RVFV strains we obtained from mosquitoes, cattle, and humans of the same area, we can reasonably conclude that these mosquito species were potential vectors for RVFV in the epizootic of 2008/2009 in the Haute Matsiatra Region of Madagascar. Clearly comprehensive studies of the three mosquito species An. squamosus, An. coustani, and Cx. antennatus, including geographical distribution, habitat preference, genetic structure of the population, vector competence, and vertical transmission of the RVFV, are essential. The knowledge gained would lead to better prevention and control of RVF outbreaks in Madagascar.
Footnotes
Acknowledgments
The authors thank the medical and veterinary staff in the districts of Fianarantsoa I, Fianarantsoa II, and Ambalavao within the Haute Matsiatra Region. The authors also thank Manfred Weidmann (Institute of Virology, University of Göttingen, Göttingen, Germany) for providing RVFV transcript RNA, Amadou A. Sall (Institut Pasteur, Dakar, Senegal) for providing the mouse RVFV immune ascitic fluid, and all the residents of the various neighborhoods for their contribution to this study. This study was supported by the Institut Pasteur de Madagascar and the Food and Agriculture Organization of the United Nations, Antananarivo, Madagascar.
Disclosure Statement
No competing financial interests exist.