
Abstract
Cattle have been recognized as hosts for Amblyomma maculatum, the Gulf Coast tick, for over 100 years. For nearly as long, A. maculatum have been known to harbor the spotted fever group Rickettsia (SFGR), now known as Rickettsia parkeri. However, human infection with R. parkeri was not documented until 2004. Results presented herein describe a laboratory and a field study evaluating cattle and the natural history of A. maculatum and R. parkeri in Mississippi. In the laboratory study, seroconversion to R. parkeri antigen occurred in calves exposed to R. parkeri by injection or by feeding R. parkeri–infected A. maculatum, and two out of six animals were transiently rickettsemic. All calves remained clinically normal during the study, except for gotch ear-like lesions in all tick-infested calves, regardless of infection status of ticks, suggesting that R. parkeri is not involved in the condition. In the field study, A. maculatum (n = 34) removed from Mississippi sale barn cattle (n = 183) and the cattle hosts were tested for R. parkeri. Cattle were not rickettsemic by polymerase chain reaction, but 49.7% demonstrated low titers to R. parkeri antigen when tested by indirect fluorescent antibody for SFGR. Of ticks removed from cattle, 11.8% were hemolymph positive and 8.7% were indirect fluorescent antibody positive. Approximately 22% (5/23) and 4% (1/23) of harvested tick extracts were positive for R. parkeri by polymerase chain reaction of the 17 kDa antigen gene and ompA gene, respectively. An amplicon for the ompA gene from one tick was successfully sequenced and showed 100% similarity with the homologous sequence of R. parkeri. Thus, cattle may harbor R. parkeri–infected A. maculatum and produce antibodies to SFGR. Cattle may play a role in the natural history of R. parkeri infection by expanding populations of A. maculatum and transporting R. parkeri–infected ticks to various locations, rather than as a reservoir for R. parkeri.
Introduction
The distribution, abundance, and occurrence of A. maculatum on wild and domestic mammals in north-central Oklahoma, and the geographical distribution of A. maculatum in Mississippi, have been well documented (Semtner and Hair 1973, Barker et al. 2004, Goddard and Paddock 2005); however, the potential role of cattle in the natural history of R. parkeri is unknown. Adult ticks prefer feeding on cattle ears, and when infestations involve sufficient numbers, the ears may become thickened and curled, causing a condition called gotch ear (Wright and Barker 2007). Little is known about the pathogenesis or epidemiology of gotch ear except that it usually involves A. maculatum (Gladney 1976, Williams et al. 1978, Byford et al. 1992, Mock 2000, Broce and Dryden 2005, Highfill 2006, Wright and Barker 2007, Wright et al. 2007).
Because infection with R. parkeri in human hosts produces a necrotic lesion (Paddock et al. 2008), we hypothesized that gotch ear might also be attributable to this infection. Further, because tick infection rates with R. parkeri appear to be higher than those with other SFGR (Sumner et al. 2007), exposure of cattle to this pathogen seems likely (Bishopp and Hixon 1936, Bishopp and Trembley 1945, Williams et al. 1978, Byford et al. 1992, Mock 2000, Barker et al. 2004, Broce and Dryden 2005, Ketchum et al. 2005, Highfill 2006, Wright et al. 2007). The aim of this study was to evaluate the role of cattle in the natural history of R. parkeri.
Materials and Methods
Clinical definition of gotch ear
We defined gotch ear as a condition of the ear in cattle associated with tick infestation, predominantly A. maculatum, exhibiting variable degrees of edema, and lesions at tick attachment sites on the outer or inner pinnae that included crusting, alopecia, erythema, and excoriation, with or without curling of the tip of the pinnae and a loss of a portion of the ear.
Experimental study in calves
All studies were approved by the Institutional Animal Care and Use Committee at Mississippi State University. Eight healthy Holstein bull-calves from the Mississippi State University dairy (age range: 3 to 6 months) with no detectable antibodies to SFGR by indirect fluorescent antibody (IFA) or detectable circulating SFGR by polymerase chain reaction (PCR) to the ompA and 17 kDa antigen genes were maintained in a tick-free environment for the entire course of the study. These were kept in biosecurity level-2 rooms located in a stand-alone building at the Mississippi State University College of Veterinary Medicine. Four calves were chosen at random for the injection group; three were inoculated by three different routes with 0.3 mL of R. parkeri–infected Vero cell suspension (Tate's Hell isolate): intradermally over the triceps muscle of the left shoulder, intravenously in the left jugular vein, and subcutaneously near the other injection sites. Growth parameters for the rickettsiae in Vero cells were as follows: inocula for all experiments were from two- to six-passage infected Vero cell suspensions with an approximate dose of 1.8 × 106 Vero cells in 1 mL for the negative control and ∼2.63 × 106 Vero cells infected with R. parkeri in 1 mL (75%–80% infected). The fourth calf served as a negative control, and was injected with 0.3 mL of an uninfected Vero cell suspension at each of the same locations.
To establish an infected tick colony, 285 engorged nymphal ticks obtained from an Oklahoma State University colony were demonstrated R. parkeri negative by hemolymph test (Gimenez 1964, Burgdorfer 1970), PCR for the ompA gene, and IFA techniques. These ticks were injected percutaneously with either R. parkeri–infected cell suspensions (n = 210 ticks) or sterile phosphate-buffered saline (PBS; pH 7.4; n = 75 ticks) using a 30-gauge needle as previously described (Goddard 2003).
All ticks were kept in vials in humidity chambers and examined daily to determine whether they had molted. In the injected tick colony, 95% (199/210) of the R. parkeri–injected ticks molted and 96% (72/75) of the PBS-injected ticks molted. The mean days to molt for the 199 ticks were 24.46 and 25.28 for the R. parkeri–injected and PBS-injected ticks, respectively. After molting to the adult stage, a subsample of 15/18 (83.3%) R. parkeri–injected ticks were positive when tested by nested PCR for the ompA gene, whereas all PBS-injected ticks were negative for this gene. The same subsample was 86% (16/18) hemolymph positive.
For the IFA test, tick hemolymph was spotted onto the wells of slides and stored at −20°C until ready to stain. A separate antigen-coated slide was used as a positive control and hemolymph-negative ticks were included as negative controls. Each sample and control field was loaded with 40 μL of anti–R. rickettsii human serum (kindly provided by Dr. William Nicholson, Centers for Disease Control and Prevention) diluted 1/100 in PBS and incubated at 37°C for 35 min. Slides were washed three times for 5 min each, twice in PBS and once in dH2O, and allowed to air-dry. Forty microliters of 1:20 fluorescein-isothiocyanate-conjugated anti-human immunoglobulin G (IgG) was pipetted onto each field and the slides were incubated again at 37°C for 35 min. The washes were repeated and the last dH2O wash was counterstained with Eriochrome black T. After drying, the slides were covered with two drops VECTASHIELD® (Vector Laboratories) and cover slips were placed over them. Sera showing titers ≥32 were titrated by twofold dilutions to their endpoints.
A second group of four calves were selected at random for the tick infestation portion of the study. Fifteen to 20 R. parkeri–infected adult A. maculatum (∼50% male) ticks were placed on the right ear of each of three calves. Similarly, 20 PBS-injected ticks were placed on the right ear of the control calf. The tick-infested ear of each calf was covered with a sock, which was adhered to the ear base with tissue glue. All ticks had either fed to repletion and remained attached, or had already dropped off by the seventh day after placement. Calves with ticks were sedated with xylazine and ticks from their ears were removed. Tick attachment sites were biopsied after local anesthesia had been applied while the calves were still under sedation from the zylazine.
At least 3 mL of whole blood in EDTA tubes and 3 mL of blood in serum separator tubes was collected from all calves once before the experiment and three times weekly after placement of ticks or injection, by jugular venipuncture for hematologic, molecular, and serologic tests. The calves were given a physical examination including monitoring of temperature, appetite, and attitude on each sampling day for 30 days. The calves' tick attachment sites were also visually inspected for signs of gotch ear. Calf sera in both groups were screened for antibodies to SFGR using R. parkeri–coated slides for IFA to detect IgG antibodies at a 1:32 dilution. Fluorescein-isothiocyanate-labeled goat anti-bovine IgG (Kirkegaard and Perry Laboratories) was used to determine serologic response to infection. Sera showing titers ≥32 were titrated by twofold dilutions to their endpoints. Anti–R. rickettsii antibodies in human serum were used for a positive control and cattle serum negative by PCR of the ompA gene and determined seronegative before the study was used as a negative control.
Skin biopsy specimens collected using an 8-mm punch were obtained from the site of tick attachment from the four tick-infested calves 7 days after exposure to R. parkeri–infected ticks as described above. Skin biopsy specimens were collected in a similar fashion (local anesthesia) from the four injected calves. Samples from all eight calves were fixed in 10% formalin and processed for histopathologic evaluation. Sections (3 μm thick) were cut from formalin-fixed, paraffin-embedded tissues and tested using an immunoalkaline phosphatase staining method for SFGR, as described in Paddock et al. (2004). DNA was extracted from blood using an Illustra Blood Genomic Prep Mini Spin kit (GE Healthcare) for all sampled days postinfection (DPI) for each calf, and the extracts were evaluated using a nested PCR assay designed to amplify a segment of the ompA gene based on a published protocol (Sumner et al. 2007). Extracts were also evaluated using a nested PCR assay designed to amplify a segment of the 17 kDa antigen gene based on a published protocol (Paddock et al. 2004). DNA extracted from cultivated Tate's Hell R. parkeri in Vero cells (CDC) was used for a positive control and water was used for a negative control. We observed the primary product by electrophoresis in 2% agarose gels containing ethidium bromide. DNA extractions, primary and secondary PCR assays, as well as gel electrophoresis were performed in separate designated areas of the lab to minimize chance of DNA contamination.
Observational study in sale barn cattle
From July through October 2008, A. maculatum ticks and whole-blood samples were collected from cattle (Table 1) at cattle auctions in six Mississippi counties while accompanying the veterinarian designated for these sales and in accordance with the Mississippi Department of Agriculture guidelines (Fig. 1). Cattle were sampled randomly, without regard to age, gender, or breed. Blood specimens were obtained from all animals regardless of the presence of ticks. DNA was extracted as described above from whole blood and samples were evaluated using nested PCR assays designed to amplify segments of the 17 kDa antigen gene and the ompA genes.
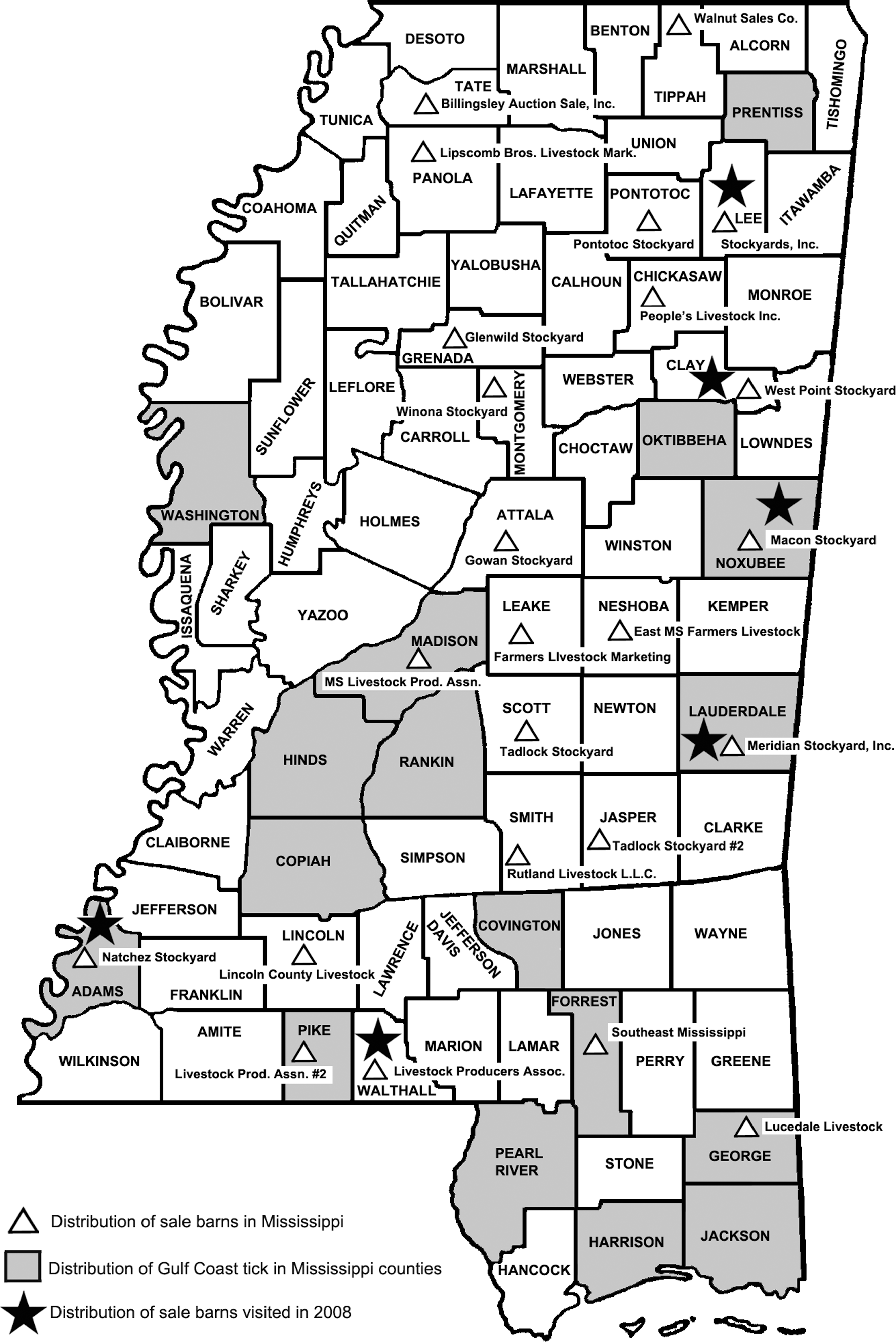
Geographic distribution of Amblyomma maculatum collections in Mississippi by county, 2005, distribution of sale barns, and distribution of sale barns visited in this study, 2008 (map constructed by Joe MacGown).
Sequence analysis for ompA amplicons showed 100% similarity with the homologous sequence of R. parkeri (GenBank accession no. U43802).
Common name for any unknown black and white cattle breed.
Ticks lost to follow-up: 5 died before hemolymph testing; another 11 died before PCR or IFA testing.
IFA, indirect fluorescent antibody; PCR, polymerase chain reaction.
Ticks collected from cattle were deposited in labeled vials for transport to the lab, where they were held in a humidity chamber until testing. Hemolymph tests (Gimenez 1964), IFA, and PCR methods were used to assay for SFGR as described above. For PCR testing, the ticks were minced using a scalpel blade and DNA was extracted using an Illustra Tissue and Cells Genomic Prep Mini Spin kit (GE Healthcare) and tested by PCR assays designed to amplify segments of the 17 kDa antigen and ompA genes. PCR products were sequenced through MWG Biotech and analyzed using ClustalX2 and the BLAST programs to identify the product (version 2.0; National Center for Biotechnology Information).
Results
Experimental study in calves
Calves exposed to R. parkeri remained alert, nonfebrile, and maintained normal appetites during the course of the study. Both negative control calves were negative for R. parkeri infection by PCR throughout the study. All calves (negative control calves and R. parkeri–exposed calves) were negative for R. parkeri using nested PCR of the ompA gene. However, two of six calves exposed to R. parkeri were transiently positive to SFGR by PCR of the 17 kDa antigen gene (one tick-infected calf on DPI-23 and one inoculum-infected calf on DPI-11 and DPI-14). Each of the six calves, exposed to R. parkeri by needle inoculation or by feeding of infected ticks, developed low IgG titers (≤64) to R. parkeri (Fig. 2) and both negative control calves remained seronegative. Attachment sites from all four calves receiving ticks showed edema, crusting and erythematous lesions on the outer pinna, and curling of the tip of the ear (Fig. 3A) and, in some cases, severe excoriations (Fig. 3B). These lesions were present in the right ear of all tick-infested calves, regardless of whether ticks were R. parkeri infected. Indurated swellings ranging from ∼2 to 6 cm developed at injection sites in all R. parkeri–injected calves as well as the control calf. Immunohistochemical staining of tissue samples collected from ears of three calves exposed to R. parkeri via tick bite and from skin at the injection site on the neck of three calves exposed via needle inoculation revealed SFGR in the inflammatory cell infiltrates (Fig. 4).
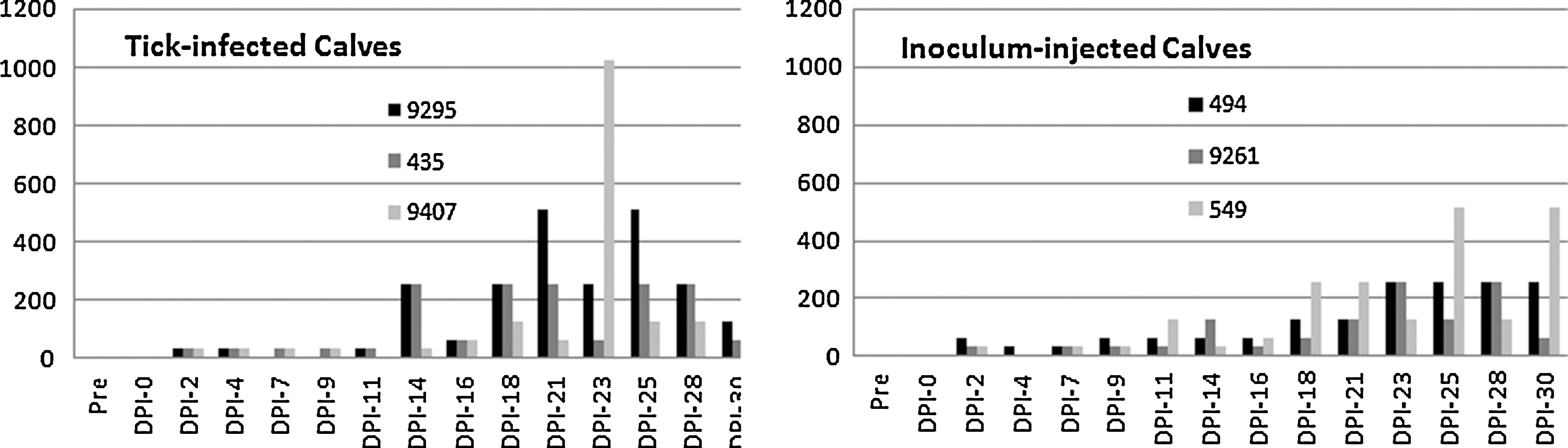
Tick-infected calves and Rickettsia parkeri–injected calves seroconverted at a 1:32 dilution on days postinfection (DPI)-2; most were transiently infected and then remained positive for the duration of the study.
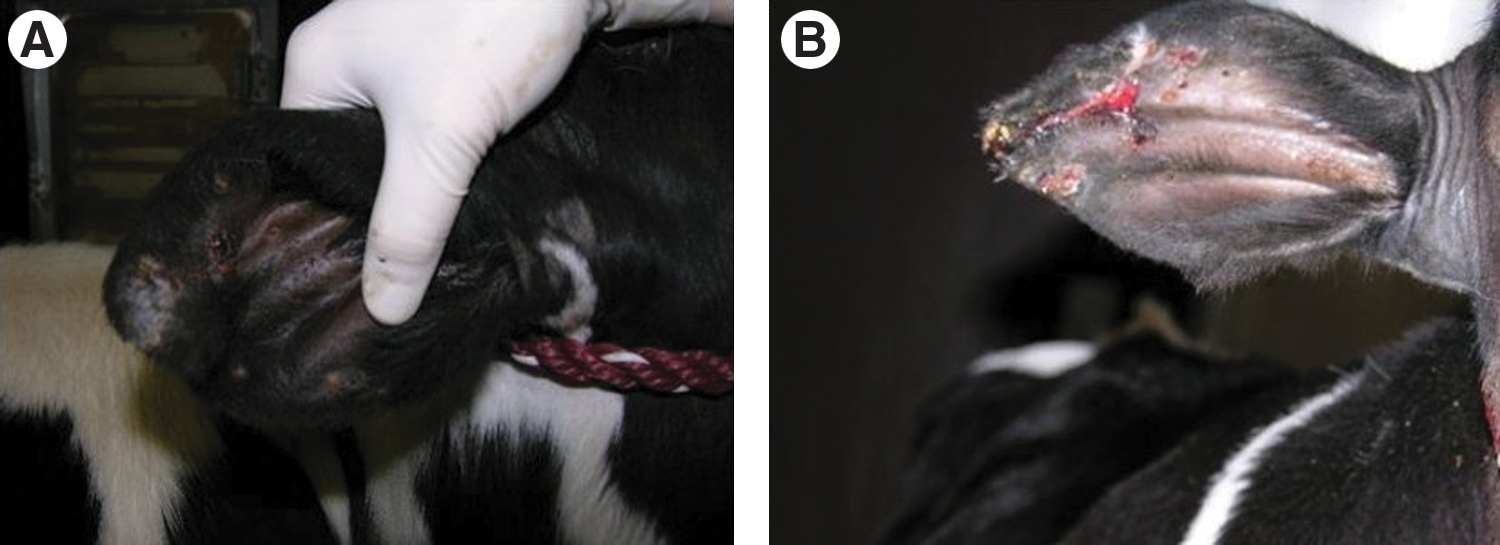
Gotch ear in calf infested with R. parkeri–infected A. maculatum ticks (
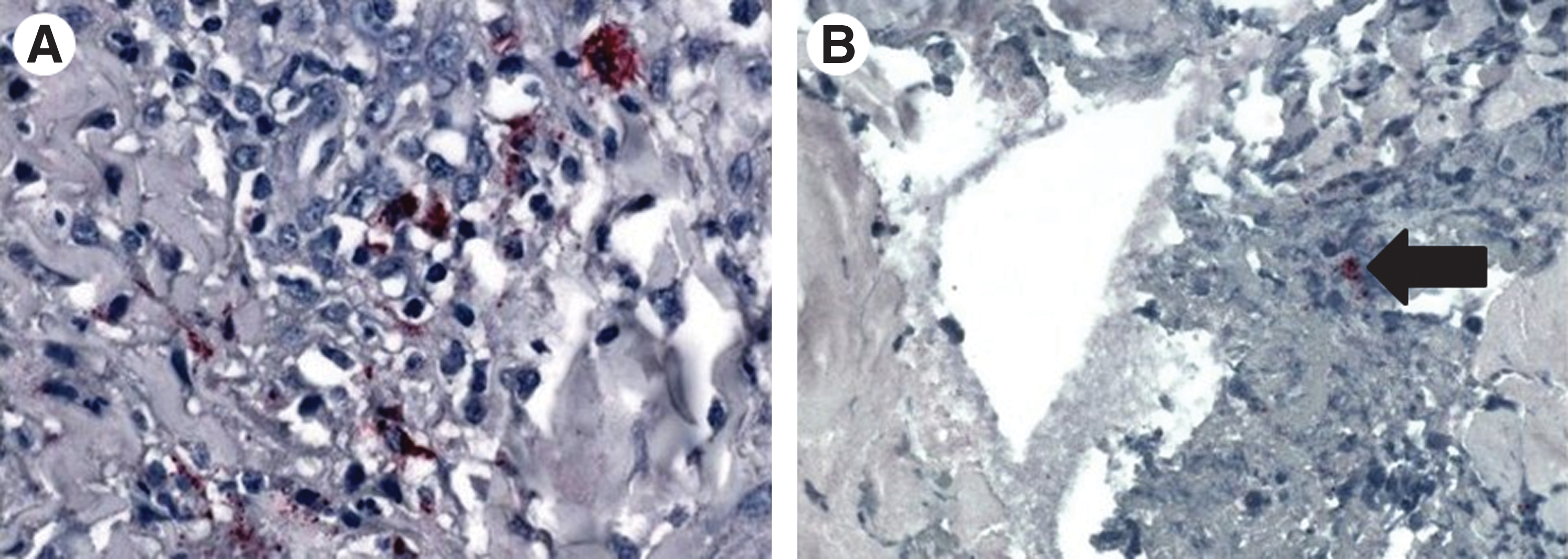
Immunohistochemistry of biopsied ear in calf infested with R. parkeri–infected A. maculatum ticks revealing spotted fever group rickettsiae in the inflammatory cell infiltrates (
Observational study in sale barn cattle
Blood samples were collected from 183 cattle at six sale barns, representing 25% of all sale barns in the state. Thirty-nine A. maculatum were removed from 13 (7.1%) cattle at 5 sale barns (Table 1). Cattle comprised a variety of beef breeds and ranged in age from 1 to 14 years (mean = 5); 176 (96%) of these animals were female. All cattle were negative for R. parkeri by PCR of both the 17 kDa antigen gene and the ompA gene. Ninety-one (50%) of cattle sera showed an IFA titer of 32 when reacted with R. parkeri antigen; however, when titrated, all samples were negative at a 1:64 dilution.
Bacteria stained by the Gimenez technique were detected in the hemolymph from 4 (11.8%) of 34 A. maculatum removed from cattle, and 2 of these also stained by IFA assay, including ticks from cattle at one sale barn in Walthall County where A. maculatum had not previously been reported (Goddard and Paddock 2005). Rickettsial DNA was detected using the 17 kDa PCR assay in 5 (22%) of 23 ticks removed from cattle at Noxubee and Lauderdale county sale barns. A 540-bp segment of the ompA gene was amplified from one of these ticks and showed 100% identity with the homologous sequence of R. parkeri (GenBank accession no. U43802).
Discussion
In this investigation, A. maculatum successfully fed to repletion on the ears of calves, and exposed calves showed a transient rickettsemia and antibody production. A cutaneous infection was identified by immunohistochemical stain in some of these calves; however, none of the R. parkeri–infected animals developed signs of systemic illness, and gotch ear occurred in all animals exposed to ticks, regardless of infection with R. parkeri.
We found no evidence of rickettsemia by PCR in any cattle, despite the relatively frequent occurrence of A. maculatum on cattle from sale barns in Mississippi, and the identification of low antibody titers to SFGR in ∼50% of these animals. However, this assessment took place at one point in time, so sera were only evaluated as either positive or negative for a particular dilution.
Using the 1:32 dilution, some experimentally infected calves (from both injected and tick-infested groups) showed an antibody response as early as DPI-2. It is difficult to explain these results, but it is noteworthy that both negative control calves were negative by IFA at both the 1:32 and the 1:64 dilutions in a blinded evaluation. A study of related rickettsiae (Rickettsia conorii) in cattle showed that the rate of seroconversion is dose related (Kelly et al. 1991). In that study, all of the cattle were inoculated with either 3000 organisms (low dose) or 100,000 organisms (high dose). The high-dose cattle showed an IgG response (titer ≥1:40) by DPI-7 and low-dose cattle by DPI-15. The dose used in our study was higher than their higher dose by a factor of nearly 30.
These data show that R. parkeri–infected A. maculatum are capable of transmitting this SFGR to naïve calves and that R. parkeri–infected ticks can be found on cattle in nature. In addition, at least some sale barn cattle demonstrated low titers of SFGR antibodies, suggesting past exposure to SFGR, possibly through the bite of infected A. maculatum.
Although there was no evidence of persistent systemic infection in cattle by R. parkeri and thus cattle do not likely serve as competent reservoir hosts for R. parkeri, we believe that cattle may play an important role in the maintenance of the rickettsia by providing a blood meal to large numbers of ticks and increasing tick populations and distribution in the southeastern United States. This may ultimately increase exposure of wildlife and people to R. parkeri–infected ticks. These results may have important implications in the development, prevention, and management approaches for gotch ear with emphasis placed on tick control rather than antibiotic treatment. Additional studies are underway in our lab to assess the role of wildlife as potential reservoir hosts for R. parkeri.
Footnotes
Acknowledgments
The authors would like to thank LARAC personnel for their kind attention to animal care throughout this study. The authors would also like to thank Jesse Carter for his skilled assistance in handling the calves. This article has been approved for publication as Journal Article No J-11653 of the Mississippi Agricultural and Forestry Experiment Station, Mississippi State University.
Disclosure Statement
None of the authors have any commercial associations that might create a conflict of interest in connection with this article. No competing financial interests exist. The findings and conclusions described in this article are those of the authors and do not necessarily represent the opinions of the U.S. Department of Health and Human Services.