
Abstract
Anopheles coustani s.l. and Anopheles squamosus are sub-Saharan mosquito species that have been implicated in malaria transmission. Although generally believed to be of negligible importance due to their overwhelmingly zoophilic behavior, An. coustani s.l. and An. squamosus made up a large proportion of the anophelines collected by human landing catches during the 2007–2008 and 2008–2009 rainy seasons in Macha, Zambia. Further, polymerase chain reaction-based blood meal identification showed that the majority of blood meals from these mosquito species caught in human-baited Centers for Disease Control light traps were from human hosts. Although no An. coustani s.l. or An. squamosus were found to be positive for Plasmodium, the demonstrated anthropophilic tendencies of these mosquitoes in southern Zambia suggest their potential as secondary malaria vectors.
Introduction
Many of the potential secondary vectors mentioned above are generally thought to be of negligible importance due to their demonstrated zoophilic behavior (Gillies and DeMeillon 1968). In particular, An. squamosus has been shown not to bite humans in the presence of alternate hosts (Symes 1931, Bruce-Chwatt et al. 1960, Hamon et al. 1964, Jupp et al. 1980), and the human blood indices (HBIs) determined for An. coustani s.l. have been low (Adugna and Petros 1996), with the majority of blood meals identified having been of bovine origin (Antonio-Nkondjio et al. 2006, Muriu et al. 2008).
In contrast to these findings, An. coustani s.l. and An. squamosus in Macha have demonstrated unexpectedly high anthropophily. An. coustani complex mosquitoes have been found harboring sporozoites in Katanga in the Democratic Republic of the Congo (Vincke 1946) and in Tanzania (Gillies 1964). Additionally, An. coustani s.l. has been found positive for Plasmodium malariae in Cameroon by enzyme-linked immunosorbent assay (ELISA) for the circumsporozoite protein (CSP) (Antonio-Nkondjio et al. 2006). Dissections of An. squamosus have found sporozoite-positive specimens in Tanzania (Gillies 1964) and Zimbabwe (Gillies and DeMeillon 1968). Because of this apparent capacity to transmit Plasmodium, the high degree of anthropophily of An. coustani s.l. and An. squamosus described here increases their importance as potential secondary vectors.
Materials and Methods
Mosquito collection and handling
Mosquitoes were collected for this study from three village areas, Chidakwa, Lupata, and Namwalinda, in the Southern Province of Zambia. Each village area is situated within 10 km of the Johns Hopkins Malaria Research Institute's field station in Macha, Zambia (16.39292S, 26.79061E) at an elevation of ∼1100 m. Collections were performed December through May during the 2007–2008 and 2008–2009 rainy seasons.
Mosquito foraging behavior, that is, the combination of host-seeking and blood feeding activity, was examined by paired indoor/outdoor human landing catch (HLC) and cattle-baited trap (CBT) collections, as well as corresponding Centers for Disease Control (CDC) light trap collections. Indoor/outdoor HLC and CBT collections were performed simultaneously on given nights at three different households, and corresponding CDC trap collections were performed on the following nights in the same sleeping houses as the HLCs. All collections were conducted for a total of 212 trap nights over the two seasons. Teams of two trained field personnel performed HLC collections from 20:00 to 06:00 during the 2007–2008 season and from 19:00 to 07:00 during the 2008–2009 season. Hourly collections were kept in separately labeled paper cups. CBT and CDC trap collections were performed for the same lengths of time as the HLCs. CBTs, kraals each big enough to encircle one calf, were constructed at each of seven collection households. The kraals were made of felled trees and branches in the style of the larger cattle kraals frequently constructed in the region. Once a calf was led into a kraal, a double-bed-sized untreated bed net was hung above the kraal so that the net rested on the outside of the enclosure, ending about 30 cm above the ground. The top of the net was loosely tied into a funnel. In the morning the upper portion of the net containing most of the resting mosquitoes was cinched closed, the calf was removed, and any remaining mosquitoes were collected by manual aspiration (Fornadel et al. in press). CDC traps were suspended next to occupied beds protected by a bed net. If the trapping room had more than one bed, the other occupants were instructed to use bed nets as well; if they had none, additional nets were provided for the night. In addition to the above catches, extra CDC light trap collections were conducted at roughly 10 households in each village area multiple times per month. Blood-fed mosquitoes from these traps were used to determine species-specific HBIs.
All mosquitoes were killed by freezing and were identified morphologically (Gillies and Coetzee 1987) at the field station. They were sorted and counted according to species and abdominal status, as well as date and location of collection. Specimens were then placed in tubes containing silica gel desiccant (Fisher Scientific) and cotton for stable storage until they were processed for molecular analysis.
DNA isolation and polymerase chain reaction
Heads/thoraces and abdomens were separated and independently rehydrated at room temperature in 20 μL of double-distilled water for 10 min before homogenization. DNA was extracted from mosquito heads/thoraces and abdomens by a modified salt procedure as described previously (Kent and Norris 2005). DNA pellets were resuspended in 50 μL of double-distilled water. For Anopheles gambiae complex mosquitoes, species was determined using the Scott et al. (1993) polymerase chain reaction (PCR) diagnostic, whereas species in the An. funestus and Anopheles longipalpis species groups were differentiated using ITS2-based diagnostics (Koekemoer et al. 2002) with An. longipalpis type C producing two ITS2 amplification products (Kent et al. 2006). The host source of blood-fed mosquitoes was determined by PCR diagnostic on abdominal DNA extractions (Kent and Norris 2005, Fornadel and Norris 2008). Plasmodium infection status was ascertained by PCR on head/thorax DNA using a novel set of primers designed to amplify a 183 bp fragment of the P. falciparum cytochrome b gene.
Primers were manually selected for the new P. falciparum diagnostic (PFcytblongF: 5′-ATACATGCACGCAACAGG TGCTTCTC-3′; PFcytblongR: 5′-CAATAACTCATTTGAC CCCATGGTAAGAC-3′) and were checked for cross-reactivity with cytochrome b sequences of An. gambiae s.s. [NC_002084.1] and An. funestus s.s. [NC_008070.1], in addition to potential host species: Homo sapiens [NC_012920.1], Bos taurus [NC_006853.1], Canis lupus familiaris [NC_002008.4], and Capra hircus [NC_005044.1]. Further, primers were experimentally tested for cross-reactivity against An. arabiensis, Anopheles quadriannulatus species A, An. coustani s.l., An. squamosus, An. longipalpis type C, and An. rivulorum. The PCR began with an initial 95°C denaturation, followed by 60 cycles of 95°C for 30 s, 58°C for 50 s, 72°C for 40 s, and a 5 min 72°C final extension. The 25 μL reactions contained 1× PCR buffer (10 mM Tris, pH 8.3, 50 mM KCl, 1.5 mM MgCl2, and 0.01% gelatin), 150 μM of each dNTP, 50 pmol of each primer, and 2 units of Taq polymerase.
The diagnostic was validated on DNA extracted as above from Zambian field samples of An. funestus, as well as from experimentally infected colonized An. gambiae s.s. (Keele strain) maintained at the Johns Hopkins Bloomberg School of Public Health. Infection status of colonized mosquitoes was ascertained by salivary gland dissection. The PFcytblong PCR was run side by side with a commonly used nested Plasmodium PCR assay (Snounou et al. 1993). About 2 μL of extracted head/thorax DNA was used as template for both reactions. The detection threshold of the primers was tested using cultured P. falciparum blood-stage parasites serially diluted with phosphate-buffered saline (104, 103, 102, 10, and 1 parasites/μL). Parasite DNA was extracted using a DNeasy Blood and Tissue Kit (Qiagen Sciences). Both the Snounou et al. (1993) and PFcytblong PCRs used 1 μL of culture DNA as template (equivalent to 104, 103, 102, 10, and 1 parasite per PCR). The PFcytblong product was easily detectable down to 1 parasite per reaction. The Snounou et al. (1993) assay was less consistent, variously detecting template out to 1 or 10 parasites.
Data analysis
As in previous studies, overdispersed mosquito count collections were modeled with a negative binomial regression analysis (Hii et al. 2000, Mathenge et al. 2004, 2005) using STATA 10 (STATACORP). The sampling efficiency of a CBT relative to an outdoor HLC pair was evaluated (Fornadel et al. in press). Briefly, mosquito counts were assumed to follow a Poisson distribution with an overdispersion parameter. Catches were modeled as a function of sampling method, collection season, and collection month. The log of the expected counts (E) was modeled as the function log(E [Yijk
]) = α +β
Results
Anopheline foraging behavior in Macha was assessed by paired indoor/outdoor HLC, CBT, and CDC light trap collections. Overall, 2998 major vectors or potential malaria vectors were captured by the paired trapping methods: 904 An. arabiensis, 872 An. squamosus, 791 An. coustani s.l., 358 An. rufipes, 43 An. pretoriensis, and 30 An. pharoensis. Other, primarily zoophilic anopheline mosquitoes caught in significant numbers were 238 An. quadriannulatus species A and 102 An. longipalpis type C. When partitioned by collection method (Fig. 1; Table 1), all anophelines in Macha displayed the same pattern of seasonality (within the collection periods December–May), with abundances peaking in February. However, because the trapping methods were variously baited, they preferentially captured different species. CDC light traps mainly collected An. arabiensis, whereas CBTs were most likely to catch An. squamosus and An. coustani s.l. Interestingly, while the majority of the anophelines captured by indoor and outdoor HLCs were the primary vector, An. arabiensis (44%), An. coustani s.l. (32%), and An. squamosus (16%) also made up a significant portion of these collections.
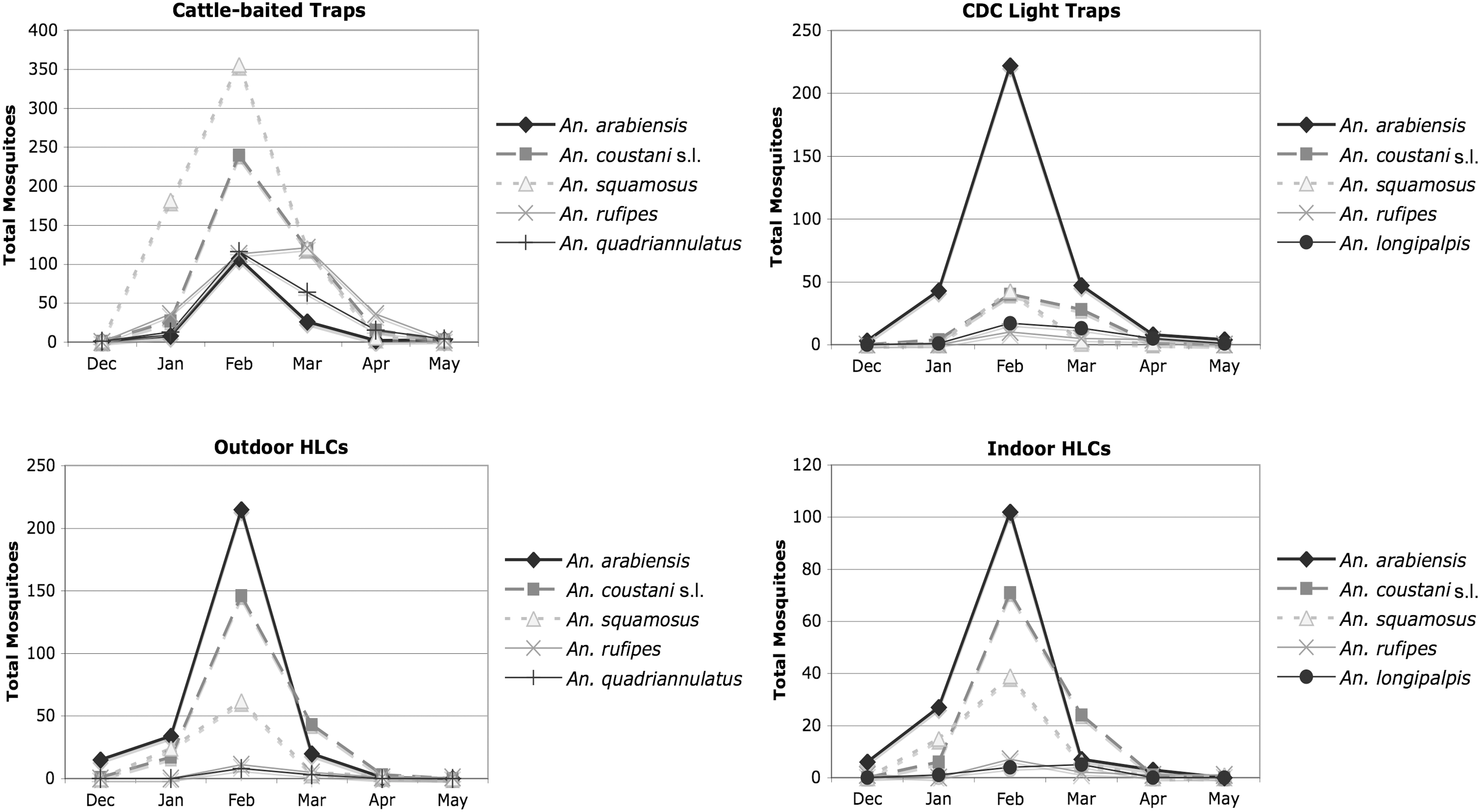
Seasonality (December–May) of anopheline mosquitoes captured during the 2007–2008 and 2008–2009 rainy seasons in Macha, Zambia. The five most numerous anopheline species captured by each method are shown. HLC, human landing catch.
Therefore, the degree of anthropophily of these potential secondary vectors was investigated further. The relative sampling efficiency of paired outdoor HLCs and CBT collections was assessed using a negative binomial regression analysis. Outside, both species displayed zoophilic behavior. CBTs captured on average 8.8 (95% confidence interval [CI]: 4.8–16.0) times as many An. squamosus and 2.2 (95% CI: 1.2–4.2) times as many An. coustani s.l. as a HLC pair during a single-trap night. However, both species were less zoophilic than the other anophelines captured in significant numbers by CBTs. An. rufipes and An. quadriannulatus were both on average 18 (95% CIs: 9–37 and 9–40, respectively) times as likely to be captured in CBTs compared with outdoor HLCs. Additionally, it was observed that An. squamosus and An. coustani s.l. readily fed on humans. About 8% of An. squamosus and 20% of An. coustani s.l. collected by CDC light trap were engorged. The HBI for An. squamosus was 0.65 and the HBI for An. coustani s.l. was 0.86 (Table 2). These high percentages were in contrast to the HBIs determined for other primarily zoophilic mosquitoes captured in CDC traps. The HBIs for An. longipalpis and An. quadriannulatus were 0.12 and 0, respectively. The majority of blood meals from these species were from cattle (Table 2). No blood-fed An. rufipes, An. pretoriensis, or An. pharoensis were collected in light traps.
Because An. squamosus and An. coustani s.l. had high HBIs, their foraging levels throughout the evening were examined (Fig. 2), since early activity would expose people to bites before they went to sleep under bed nets. Inside, An. squamosus biting behavior fluctuated throughout the night without peaking significantly, whereas outdoors, an early peak in activity was seen between 20:00 and 22:00, with another peak around midnight followed by a steady decline in activity. An. coustani s.l. was most active from 20:00 to 21:00 outdoors with its biting activity steadily declining throughout the night. An early foraging peak, although more moderate, was also seen indoors from 20:00 to 22:00.

Total Anopheles coustani s.l. and Anopheles squamosus captured each hour by indoor and outdoor HLC collections.
Over the 2 years of this study 806 An. squamosus and 303 An. coustani s.l. were screened for P. falciparum. None were found to be positive by PCR, but none of the An. arabiensis tested from the same collections were found to be positive either.
Discussion
There are seven main malaria vectors recognized on the African continent: An. gambiae s.s., An. funestus, An. arabiensis, Anopheles nili, Anopheles moucheti, Anopheles melas, and Anopheles merus (Gillies and DeMeillon 1968). A number of secondary vectors have also been recognized, including An. squamosus and An. coustani s.l. These species have been considered of negligible importance because of their strong zoophilic tendencies. However, here we report that An. coustani s.l. and An. squamosus displayed unexpectedly high anthropophily in southern Zambia, giving them a greater potential to impact malaria transmission.
Both An. coustani s.l. and An. squamosus made up a significant proportion of the anophelines captured by HLC. An. coustani s.l. made up 32% of the HLC collection in this study. This differs markedly from HLC collections made in Cameroon (Antonio-Nkondjio et al. 2006), Kenya (Mbogo et al. 1995), and Senegal (Dia et al. 2008), where the species made up <1% of the anophelines captured. However, the high degree of anthropophily in the An. coustani s.l. population in Macha is similar to the results of HLCs performed in Ethiopia, where 31.5% of the anophelines collected were An. coustani s.l. (Taye et al. 2006). Additionally, it has been reported that An. coustani s.l. readily bites man in South Africa (Coetzee 1983) and was found to make up 14% of the HLC collections in Mozambique (Mendis et al. 2000). An. squamosus made up 16% of our HLC collections. This is in contrast to findings from a number of countries. In South Africa, only one An. squamosus was captured on human bait (Jupp et al. 1980), whereas collection frequencies reported from Haute-Volta (Hamon et al. 1964), now Burkino Faso, ranged from 2.8% to 8.3% depending on the season. In addition, An. squamosus in Kenya has been shown to make up <1% of human-baited collections (Mbogo et al. 1995). Taken together, these surveys demonstrate that the foraging behavior of An. squmosus and An. coustani s.l. is regionally variable, with local anthropophilic behaviors potentially increasing the mosquitoes' roles as a secondary malaria vectors in certain areas.
To further address the anthropophily of An. squamosus and An. coustani s.l. the HBIs of these species were evaluated. The majority of blood meals from engorged specimens collected by CDC light trap in this study were from human hosts, demonstrating a high degree of anthropophily by these mosquitoes in Macha. There have been no other reports of HBIs this high for An. coustani s.l. or An. squamosus. In Cameroon (Antonio-Nkondjio et al. 2006) the only engorged An. coustani s.l. that were collected had fed on cattle, while low HBIs of 0.26 and 0.054 were observed in Ethiopia (Adugna and Petros 1996) and Kenya (Muriu et al. 2008), respectively. Reported HBIs for An. squamosus have also been low: 0.058 for specimens tested from Ghana, Nigeria, Zimbabwe, Zanzibar, and Burkina Faso (Bruce-Chwatt et al. 1960) and 0.036 from collections in Kenya (Symes 1931). In addition to a true regional variance in host preference, differences in HBIs between these studies might reflect the availability of alternate hosts.
An. squamosus and An. coustani s.l. in Macha both displayed exophagic tendencies, along with early evening foraging behavior. Similarly, An. coustani s.l. in Nigeria (Hanney 1960), Mozambique (Mendis et al. 2000), and Ethiopia (Taye et al. 2006) displayed peak biting outdoors before 21:00, and An. squamosus has shown early peaks in foraging activity followed by moderate biting throughout the remainder of the night (Van Someren et al. 1958, Hamon 1963). The combination of outdoor and early evening foraging behavior for these species could increase their potential as secondary vectors in areas where indoor control measures such as indoor residual spraying or ITNs are employed.
Interestingly, during the 2 years of this study only two An. pharoensis were captured by HLC, whereas 24 were caught in CBTs. Therefore, while An. pharoensis is a recognized secondary vector in Kenya (Mukiama and Mwangi 1989), Egypt (Barber and Rice 1937), Tanzania (Draper and Smith 1957, Gillies 1964), Cameroon (Robert et al. 1992, Antonio-Nkondjio et al. 2006), Senegal (Carrara et al. 1990, Dia et al. 2008), Chad (Kerah-Hinzoumbe et al. 2009), and Mali (Holstein 1951), in southern Zambia this species most likely does not contribute to malaria transmission. The vastly different behavior of An. pharoensis in Macha might be associated with the existence of an An. pharoensis species complex, as suggested by Miles et al. (1983). Likewise, An. coustani s.s. has been shown to have a cryptic sibling species, An. crypticus, based on polytene chromosomes and cross-mating studies (Coetzee 1983, Coetzee 1994). Observed variation in anthropophilic behavior among geographically distinct populations of An. coustani s.l. could be due to the possibility of unidentified sibling species of the An. coustani complex. Likewise, An. squamosus may have a cryptic sibling species. It might also be that the observed differences in anthropophily rates of An. coustani s.l. and An. squamosus are due to higher population densities of these species, leading to a portion of the population feeding on human hosts. More work on these neglected anopheline species will be needed to explain the variation seen in their foraging behaviors between different countries across Africa.
A number of Plasmodium diagnostics have been developed to determine infection rates in mosquitoes. CSP ELISA (Burkot et al. 1984, Beier et al. 1987) is one of the most widely adopted techniques, but CSP ELISA may not pick up low level infections (Beier et al. 1988) as might be expected in potential secondary vectors. PCR-based assays that are highly sensitive do exist, but most have their own drawbacks, needing salivary gland dissection before PCR (Tassanakajon et al. 1993) or requiring multiple reactions (nested PCR) (Snounou et al. 1993) that are more prone to contamination. Therefore, we developed a single-step assay that would enable us to detect very low infection rates.
Although no Plasmodium-positive An. squamosus or An. coustani s.l. were found in Macha during the course of this study, neither were any positive An. arabiensis captured in the same collections, even though HBIs of An arabiensis were >90% during each year of this study (Fornadel et al. in press). P. falciparum was previously hyperendemic in Macha, but recently there has been a dramatic reduction in pediatric malaria admissions to the Macha Mission Hospital (Thuma 2007), most likely due to a combination of the adoption of artemesinin combination therapy as the standard treatment for uncomplicated malaria in Zambia, as well as concerted vector control efforts, including large-scale ITN distribution (Chanda et al. 2008). As malaria in the region moves toward hypoendemnicity, disease incidence has become very focal. Therefore, specimens of An. squamosus and An. coustani s.l. will need to be collected over larger areas where active transmission is ongoing so that the role of these species as potentially secondary vectors can be further evaluated.
Footnotes
Acknowledgments
We thank Vandana Singhal for her help with the infectious An. gambiae s.s. feeds and Davison Sangweme for his help with mosquito dissections. We would like to especially thank Dr. Marie Diener-West for her statistical insights, as well as Shadrack Habbanti for his time and effort spent coordinating field team operations in Zambia and Musapa Mulenga for managing our collections in Macha. We also thank our field mosquito collectors: Corrence Munsanje, Nathan Phiri, Clement Mwaanga, Fines Mwaanga, Twaambo Moono, Gift Shamapani, Miyanda Moono, Guide Hansumo, Mathias Muleka, Chaltone Munsanje, Pathias Chibambo, Malony Mulota, Haggard Musyutila, Cliff Singanga, Paul Haakaloba, and Ojukwu Himumwe. Research funding was provided to D.E.N. from the Johns Hopkins Malaria Research Institute, NIH training grant (T32AI007417) support to L.C.N., a Johns Hopkins Bloomberg School of Public Health Sommer Scholarship award to C.M.F., and additional training support from Fogarty (5D43TW001587).
Disclosure Statement
No competing financial interests exist.