
Abstract
This study aimed to investigate the occurrence of coronaviruses (CoVs) in captive birds placed inside a zoological park in Brazil. The role of captive birds in the epidemiology of CoVs in the tropics is poorly understood. A total of 25 (n = 25) different species were tested for viral RNA using individual fecal samples collected from healthy birds. Reverse transcription–polymerase chain reaction targeting the 3′ untranslated region was used to detect CoV RNA, and positive samples were submitted for sequence analysis. The phylogenetic search revealed nine mutations in the black shouldered peafowl (Pavus cristatus) CoV sequence, which clustered separately from samples previously described in England. This is the first report on the detection of the CoV genome in captive birds in Brazil.
Introduction
Viral diseases can cause substantial mortality in both wild and captive bird populations. Brazil is a country with many migrating wild birds, which is likely to facilitate disease transmission from migratory to nonmigratory flocks in zoological parks or on farms. As viral detection and identification techniques become increasingly sophisticated, and as the study of wildlife diseases increases, it is likely that new viruses will be discovered and new diseases will be encountered (Hughes et al. 2009). Therefore, studies of the viral diseases of wild and captive birds are necessary. The latest techniques of molecular epidemiology can help scientists understand how viruses move from species to species and from region to region (Hughes et al. 2009, Jackwood et al. 2010).
The present study demonstrates the molecular detection and characterization of viruses from captive exotic bird populations sampled in São Paulo State Zoological Park in Brazil, which appear to be related to group 3 CoVs.
Materials and Methods
Study design
Serial cross-sectional surveys of captive wild bird populations, maintained in a zoological park in the city of Bauru (N22°10′ S49°00′ W), were undertaken in July 2009. Feces were collected from 25 different species of healthy birds (Table 1), placed in virus transport media (Eagle's minimum essential medium supplemented with 10% fetal calf serum, penicillin, and streptomycin suspension, plus amphotericin B), and frozen at −80°C for individually analysis later. This work was authorized by the Animal Ethics Committee of the São Paulo State University (protocol number 2008/09878-09).
Ad., adult (>2 years).
Juv., juvenile (<2 years).
Viral RNA extraction
Two grams of feces was homogenized in two volumes of Eagle's essential medium (Invitrogen™; pH 7.4) and clarified by centrifugation at 3000 g for 20 min. The supernatant was filtered once through a 0.75-μm paper filter (Millipore™) and then twice through a 0.25-μm syringe filter (Corning™) as described previously (Teixeira et al. 2007). These suspensions were heated at 100°C for 30 min in a water bath before total RNA extraction was performed. Total RNA was extracted using Trizol® standard protocol, based on guanidinium isothiocyanate and acid–phenol with some modifications (Teixeira et al. 2007). We mixed 200 μL of clinical suspension with 500 μL of Trizol reagent and incubated for 10 min at room temperature. After addition of 200 μL of chloroform, the reaction was mixed vigorously for 10 s and centrifuged at 13,000 g for 10 min. The upper aqueous phase was mixed with an equal volume of cold isopropanol and incubated on ice for 10 min. The total RNA precipitate was pelleted by centrifugation at 13,000 g for 10 min and washed with ethanol. The RNA was dissolved in 30 μL of diethylpyrocarbonate-treated, sterile, double-distilled water and stored at −20°C.
Reverse transcription–polymerase chain reaction
The primer combination untranslated region (UTR) 11 − /UTR41+ , which corresponds to the conserved region of the IBV 3′ UTR and produces a very sensitive reverse transcription–polymerase chain reaction (RT-PCR) result (Adzar et al. 1997, Culver et al. 2006), was used. Before RT-PCR was performed, RNA was heated for 3 min at 100°C, followed by 2 min at 72°C, and finally for 2 min at 50°C. Optimization of a one-step RT-PCR requires compromises in the concentrations of PCR reagents, particularly the primers used. The RT-PCR was performed following the instructions in the One-Step Superscript III® RT-PCR commercial kit. Briefly, 2 μL of the first-strand product was used as a template for amplification in a 50 μL reaction mixture containing 20 pmol of primers and 1.6 mM MgCl2. All PCR conditions were the same as described above, but the amount of RT-Taq DNA polymerase platinum enzymes was calculated to 2.5 units/50 μL reaction mixture. A total of 10 μL of PCR products was electrophoresed at 100 V for 1 h using a 1.5% agarose gel in Tris-borate EDTA buffer and visualized with ethidium bromide staining and an ultraviolet transluminator. Gel images were captured using a Kodak DC290 digital camera and ADOBE 6.0 software.
DNA sequencing and phylogenetic analysis
The PCR products were purified using a commercial purification kit (QIAquick PCR purification kit; Qiagen) according to the manufacturer's instructions and sequenced directly with an ABI 3730xl DNA analyzer (Applied Biosystems). Nucleotide sequences derived from this study have been deposited in the GenBank sequence database under the following accession numbers: GU323504, Agamia/BR/UNESP/2009; GU323505, APatagonius/BR/UNESP/2009; GU323506, Pavus/BR/UNESP/2009; GU323507, Pavus/2/BR/UNESP/2009; GU323508, Penelope/BR/UNESP/2009; GU323510, Martinica/BR/UNESP/2009; GU323509, Martinica/BR/UNESP/2009. The CoV sequences AJ619580 (pheasant CoV ph/UK/602/95), AJ619579 (pheasant CoV ph/UK/438/94), AJ310642 (Turkey CoV turkey/UK/412/00), AJ278338 (IBV HVI-140), AJ278337 (IBV HV10), and AJ278335 (IBV D207) were obtained from GenBank for use in the phylogenetic analyses. Sequences were assembled using SeqMan Pro., and genome annotation was conducted with SeqBuilder (DNASTAR, Inc., v.8.0.2). Nucleotide and deduced amino acid sequences were aligned using Clustal-W in the MegAlign program (DNASTAR, Inc.). A phylogenetic tree was constructed based on amino acid alignments using BLASTN analysis (
Results and Discussion
Captive animals in zoos are part of a complex ecosystem that includes free-roaming animals and their associated infectious diseases, including respiratory viruses. These results, obtained from captive birds in Brazil, represent the first report of a circulating CoV inside a zoological park. The CoV RNA was detected in eight individual fecal samples (Table 1), which corresponds to an individual prevalence estimate of 1.12% (95% confidence interval 0.5–2.7). Of the samples considered positive, two were from black shouldered peafowl (Pavus cristatus), two were from American purple gallinules (Porhyrula martinica), and one each came from a Chestnut-bellied heron (Agamia agamia), an Emperor penguin (Aptenodytes patagonicus), a Rusted margined guan (Penelope supercilliaris), and a Roadside hawk (Rupornis magnirostris). Interestingly, all of the positive samples were obtained from adult birds. Phylogenetic analyses were based on a sequence of 146 nt after the removal of primer sites (Fig. 1). Most remarkably, nine mutations were revealed in the Pavus/2/BR/2009 (GU323507) sequence, despite many reports that consider the 3′ UTR region to be highly conserved among group 3 CoVs (Cavanagh et al. 2002). One hypothesis that could explain these mutations is the high density of different species of birds located in the same geographical area, which allows the virus to circulate among species (Jackwood et al. 2010). On the other hand, the natural habitat provides more distance between species, particularly between predators and prey. In addition, the sequences of the CoVs isolated from the two peafowl samples clustered separately from the British deposited sequences, as illustrated in Figure 1 (Cavanagh et al. 2002). Sequences within this cluster were relatively homogenous between two European samples (Fig. 1), in accordance with previous studies (Cavanagh et al. 2002).
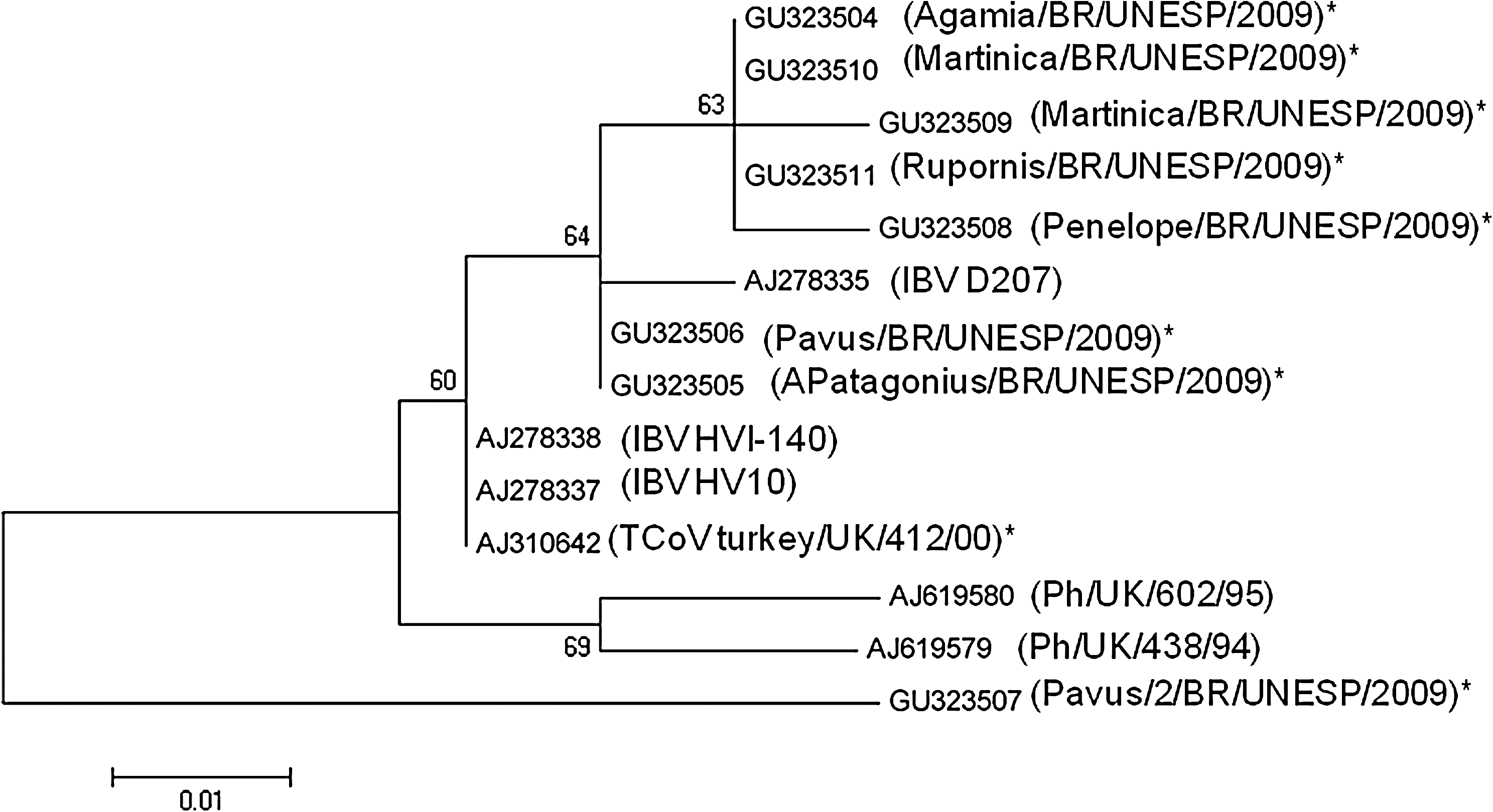
Minimum-evolution tree of coronaviruses
Samples obtained from the Chestnut-bellied heron, two American purple gallinules, a Roadside hawk, and a rusted margined guan clustered together in a single clade, close to the sequence of the IBV D 207 strain (Figs. 1 and 2). In the present study of samples collected from 25 different species (kept in isolated locations within the zoological park), CoV RNA was detected in two Galliformes, two Ciconiiformes, one Gruiforme, and one Sphenisciforme. This demonstrates the high diversity in the susceptible species, as described previously (Woo et al. 2008). In fact, CoVs have been also detected in wildfowl species, rock doves, wild peafowl, waders, Amazon parrots, and some passerine species (Cavanagh 2005, Liu et al. 2005, Gough et al. 2006, Woo et al. 2008, Hughes et al. 2009). The hypothesis that wild free-flying birds present in zoos may often act as vectors flying between enclosures that house the exotic birds and transmitting infections is not fully understood.

A comparison of part of the 3′ untranslated region from three CoVs isolated from chickens (prefixed IBV), one CoV from turkeys (prefixed TCoV), and two CoVs from pheasants (prefixed ph), with the same region identified in the eight CoVs isolated from the captive birds described in this study. Dots indicate that all the viruses had the same nucleotide at that position. Deletions are marked with an asterisk. Yellow shading indicates the mutations observed in the Pavus/2/BR/UNESP/09 sequence at a given position.
To understand the potential role of captive populations as reservoirs for CoVs, further studies are needed not only in South American zoo parks, but also worldwide. This is the first description of the molecular detection of group 3 CoVs related to captive birds in South America. The genetic diversity found in the black shouldered peafowl samples raises interesting questions about the 3′ UTR conserved region, its evolution, and viral circulation among different species inside zoo parks.
Footnotes
Acknowledgments
This work was supported by FAPESP (Fundação Amparo à Pesquisa do Estado de São Paulo) and CNPq. The authors are indebted to the technical team of the zoological park for their support. Tereza Cristina Cardoso is a recipient of CNPq grant.
Disclosure Statement
No competing interests exist.