
Abstract
West Nile virus (WNV) and Japanese encephalitis (JE) virus are distributed separately in the world with some exceptions. There is a concern that WNV may invade into Asia where JE virus exists. On and after such invasion, any differential diagnosis could be complicated by serological crossreactivities. We previously demonstrated experimentally using horses infected with WNV that preimmunization with inactivated JE vaccine considerably affected the ability of neutralization tests and immunoglobulin M (IgM) antibody-capture enzyme-linked immunosorbent assay (ELISA) to diagnose WNV infection. Here, we investigated WNV-specific antibody responses in vaccinated horses using a blocking ELISA and complement-dependent cytotoxicity (CDC) assay to evaluate these two newly developed serodiagnostic methods for WNV infection. Sera previously collected from six experimentally infected horses were used: Three were vaccinated before the infection, whereas the other three remained unvaccinated. WNV-specific antibody responses were successfully detected in the vaccinated and unvaccinated horses using both new methods, except for one vaccinated horse in which responses were not induced, probably as a result of crossprotection induced by JE vaccination. Specific antibody responses were at earliest detected from days 9 to 10 postinfection in the blocking ELISA, whereas the CDC assay provided earlier detection (at days 7–8) in all horses. The time courses of antibody levels were similar between vaccinated and unvaccinated horses in either method, indicating no notable effect of vaccination on detection of specific antibody responses, as far as antibodies were induced. These results indicated that blocking ELISA, but preferably the CDC assay, can be useful for detecting WNV infection in JE-vaccinated horses.
Introduction
The clinical manifestations of WNV infection are similar to those of JEV in humans (Solomon and Vaughn 2002) and horses (Endy and Nisalak 2002, Castillo-Olivares and Wood 2004). Therefore, differential diagnosis must rely on laboratory tests, which presently include virus isolation or viral RNA detection, along with serological tests to detect antibodies to WNV (Shi and Wong 2003, Castillo-Olivares and Wood 2004). A firm diagnosis is provided by virus isolation or viral RNA detection, but the period of viremia/RNAemia is limited, often making such a method not so useful, even at the time of illness onset (Sejvar and Marfin 2006); thus, serological testing is important. The serological tests for WNV infections recommended by the manuals and guidelines created in Japan (Kobayashi and Kurane 2003, Ministry of Agriculture, Forestry and Fisheries, Japan 2003, Kurane 2005) include a neutralization test and immunoglobulin M (IgM) antibody-capture enzyme-linked immunosorbent assay (ELISA) (MAC-ELISA), both generally recognized as specific for diagnosing flavivirus infections.
WNV is in the JE serological group, as is JEV (Mackenzie et al. 2002). The antigenic crossreactivity between WNV and JEV is problematic for differential serodiagnosis. In Japan where JEV exists, most humans and horses have been vaccinated with JE vaccine and/or naturally infected with JEV. Therefore, preimmunity against JEV is thought to induce secondary antibody responses crossreactively on infection with WNV. This was demonstrated in our earlier study using horses experimentally infected with WNV (Shirafuji et al. 2009) or by others using mice (Lim et al. 2008): Levels of neutralizing antibody responses to heterologous antigens (JEV) rapidly elevated in an anamnestic fashion were equivalent to or even higher than those to homologous antigens (WNV) produced by the primary antibody responses. This complication also occurred in a MAC-ELISA in which antibody responses to WNV infection were infrequently detected in horses preimmune to JEV (Shirafuji et al. 2009).
Our laboratory has developed an epitope-blocking ELISA and a complement-dependent cytotoxicity (CDC) assay for detecting antibody responses specific for WNV, but not crossreactive to JEV in horse sera (Kitai et al. 2007, 2010). The blocking ELISA is based on the binding of a specific antibody to the antigen that is inhibited by its preceding incubation with a test antibody specimen (Kitai et al. 2007). The CDC assay utilizes a mechanism of antibody-mediated CDC in which complement activation triggered by a specific antigen-antibody binding on the cell surface causes cell lysis (Kitai et al. 2010). For these assays, we used the nonstructural protein 1 (NS1) of WNV as an antigen: We generated a monoclonal antibody specific for the NS1 of WNV and also a cell line stably expressing the NS1 antigen of WNV on the cell surface and in a secreted form (Kitai et al. 2007). Both assay methods successfully detected antibodies reactive to WNV but not to JEV. However, horses used for these experiments (Kitai et al. 2007, 2010) were naïve, representing primary infection with WNV but not adapting to the situation in Japan where almost all horses are immune to JEV.
The present study aimed at investigating WNV-specific antibody responses in horses vaccinated with inactivated JE vaccine and then experimentally infected with WNV. WNV-specific antibody responses detected by the blocking ELISA and CDC assay were compared with those detected by conventional serodiagnostic methods to evaluate the new assays. The results obtained in the present study indicate the usefulness of both new methods. However, the CDC assay appears preferable for diagnosis of WNV infection.
Materials and Methods
Cells
Cell lines stably transfected with NS1 (2G2 cells) or NS1/NS2A genes (2G12 cells) of the WNV Eg101 strain have been described (Kitai et al. 2007). Both cells secrete NS1 antigen. Culture fluids of 2G2 or 2G12 cells were used as the antigen for the conventional ELISA for measuring antibodies to NS1 or for the blocking ELISA, respectively. For the CDC assay, 2G2 cells were used as the antigen.
Antibodies
Monoclonal WN-2H4 specific for the NS1 of WNV has been described (Kitai et al. 2007). The monoclonal was obtained in ascites form from pristane-primed BALB/c mice. Rabbit anti-NS1 hyperimmune serum was obtained in our earlier study (Kitai et al. 2007) by repeated immunization of a Japanese white rabbit with the NS1 antigen affinity purified from culture fluids of WNV-infected Vero cells.
Horse sera
All horse sera used in the present study were those obtained and used in our previous study (Shirafuji et al. 2009). Briefly, six 1- to 2-year-old horses (horses #1–#6) were used. Three horses (#s 4, 5, and 6) were vaccinated twice with a 4-week interval by intramuscular inoculation with inactivated JE vaccine (BM III strain; Nisseiken, Tokyo, Japan). Four weeks after the second vaccination, these horses, as well as the other three unimmunized ones, were infected subcutaneously with 1 × 107 PFU of the NY99 strain of WNV. After infection, these horses were bled at 1–10, 14, and 21 days postinoculation. For the CDC assay, sera were heat inactivated at 56°C for 30 min before use.
Conventional ELISA to measure NS1 antibody levels
NS1 antibody levels in horse sera were essentially measured following a conventional ELISA as previously described (Kitai et al. 2007). Briefly, microplates sensitized with 3 ng of NS1 antigen per well were incubated serially with test sera of 1:100 dilution, alkaline phosphatase-conjugated rabbit anti-horse IgG, and p-nitrophenyl phosphate. The NS1 antigen was affinity purified from culture fluids of 2G2 cells by a monoclonal WN-2H4. To minimize nonspecific reactions, a nonsensitized control plate was run in parallel, and the difference in absorbances from antigen-sensitized wells was obtained.
Blocking ELISA
The blocking ELISA was performed as previously described (Kitai et al. 2007). Briefly, microplates were incubated serially in the following steps: (1) rabbit anti-NS1 hyperimmune serum; (2) culture fluids of 2G12 cells; (3) test sera; (4) WN-2H4; (5) alkaline phosphatase-conjugated goat anti-mouse IgG; and (6) p-nitrophenyl phosphate. Test sera were incubated in parallel with the ELISA diluent in step (3), and WN-2H4 (subclass; IgG1) with mouse IgG1 (without any anti-WNV activity) in step (4). The reason for the use of mouse IgG1 in step (4) is to minimize a weak reactivity that the rabbit anti WNV NS1 hyperimmune serum used for the capture antibody against mouse IgG1. The percentage inhibition of monoclonal antibody binding was calculated from absorbances by the formula 100−100 × (A−B)/(C−D), where A is the absorbance obtained combining steps (3) and (4) with test sera and WN-2H4, B is obtained with test sera and purified IgG1, C is obtained with ELISA diluent and WN-2H4, and D is obtained with ELISA diluent and purified IgG1. The cutoff value to determine the presence of WNV-specific NS1 antibodies in test sera was 27.6% of the inhibition in the blocking ELISA.
CDC assay
The CDC assay was performed as previously described (Kitai et al. 2010). Briefly, 50 μL of heat-inactivated test serum was mixed with an equal volume of 2G2 cell suspension containing 5 × 104 cells and incubated on ice for 30 min. The mixture was then mixed with 11 μL of rabbit complement to make a final concentration of 10% and incubated at 37°C for 2 h. After centrifugation, 50 μL of the supernatant was mixed with 50 μL of a lactose dehydrogenase substrate (Cytotoxicity Detection Kit Plus [LDH]; Roche, Mannheim, Germany) and incubated at room temperature for 15 min, followed by spectrophotometry at 490 nm. The percentage of specific cell lysis was calculated using the following formula: 100 × [(A−C)/(B−C)], where A represents an absorbance obtained with test serum, B represents an absorbance obtained by lysing all the target cells with 1% Triton X-100, and C represents an absorbance obtained with target cells incubated in medium containing rabbit complement at 10%. When this calculation provided a negative value, 0.0% was assigned to the result. The cutoff value to show the presence of WNV-specific NS1 antibodies in test sera was determined as 19.8% of specific lysis in the CDC assay (Kitai et al. 2010). In the one-dilution method, the percentage of specific cell lysis obtained at a 1:10 dilution of sera was used as the NS1 antibody level. In the endpoint method, the NS1 antibody titer was expressed as the highest serum dilution giving >19.8% specific lysis.
Results
Since the present study used horse sera obtained in an earlier study of ours (Shirafuji et al. 2009), levels of viral RNA, neutralizing antibody titers, IgM antibody levels, and clinical manifestations were known. These results are shown in Figure 1, along with those obtained in the present study.
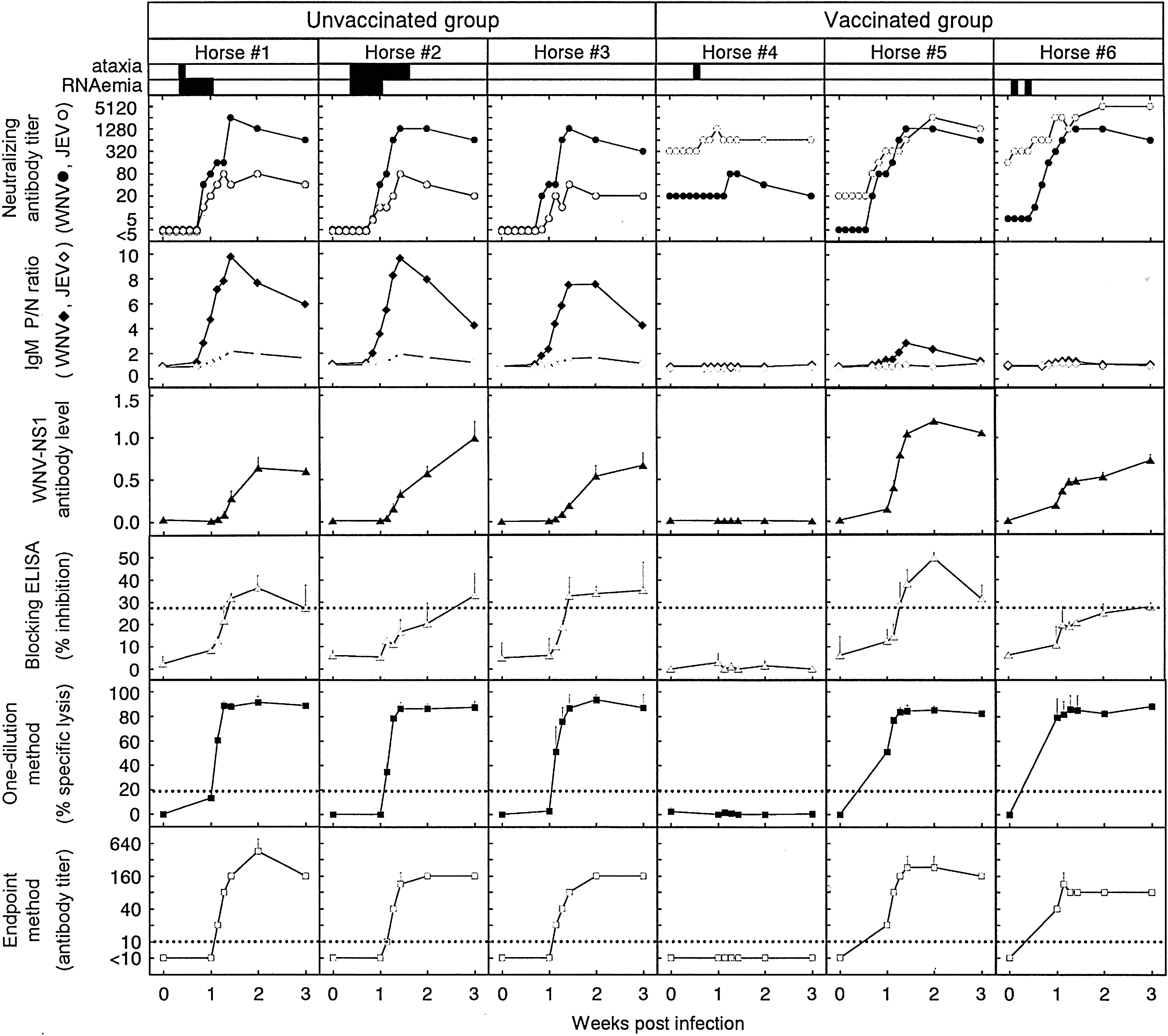
Time courses of antibody responses to West Nile virus (WNV) in individual horses experimentally infected with WNV. Results obtained in the present conventional and blocking enzyme-linked immunosorbent assays (ELISAs) and complement-dependent cytotoxicity (CDC) assay are shown along with those obtained in an earlier study of ours (Shirafuji et al. 2009) by a neutralization test and MAC-ELISA, as well as development of clinical manifestation (ataxia) and RNAemia. Sera were serially collected from six horses inoculated subcutaneously with 1 × 107 PFU of the NY99 strain of WNV: horses #4–6 were vaccinated with inactivated Japanese encephalitis (JE) vaccine before infection. These sera were tested by the TaqMan RT-polymerase chain reaction assay for the presence of WNV RNA, neutralization test against WNV (closed circle) or JE virus (JEV) (open circle), MAC-ELISA for immunoglobulin M antibodies to WNV (closed diamond) or JEV (open diamond), conventional ELISA to measure antibodies to WNV NS1, blocking ELISA, and a one-point method and an endpoint method of the CDC assay. Results of MAC-ELISA were represented as positive/negative (P/N) ratios. Each datum obtained with conventional ELISA, blocking ELISA, and CDC assay represents an average obtained from two separate experiments (standard deviations are indicated by bars). Dotted lines indicate the cutoff value in the blocking ELISA (27.6%) and CDC assay (19.8% for the one-point method, 1:10 for the endpoint method). The presence of ataxia and RNAemia were monitored each day and are indicated by closed bars.
ELISA antibody levels to NS1
The neutralization test and MAC-ELISA measure antibodies to E, a structural protein of WNV. Since the blocking ELISA and CDC assay detect antibody responses to NS1, a nonstructural protein, a conventional ELISA was used to confirm the induction of antibody responses to NS1 in WNV-infected horses before evaluating these two new assays. Since JEV NS1 is not included in the JE vaccine preparation (Konishi et al. 2004), the antibody levels shown by the conventional ELISA represent specific but not crossreactive responses to WNV NS1. As shown in Figure 1, all but one (horse #4) of the six horses developed NS1 antibodies at levels of 0.593–1.049 at 21 days after infection. NS1 antibody levels started to increase on day 8 in the three horses that did not receive vaccination (horses #1–3), whereas NS1 antibody levels started to increase by day 7 in horses #5–6.
Inhibition values in blocking ELISA
Horses with or without vaccination were examined for antibody responses to WNV infection using the blocking ELISA. The percent inhibition of WN-2H4 binding was compared between horses vaccinated and unvaccinated before WNV infection (Fig. 1). In all three unvaccinated horses, WNV-specific NS1 antibody responses were detected at 21 days after infection at the latest: The earliest detection was on day 10 in horses #1 and #3. The inhibition values for these three horses were up to 36.5%, not as high as those reported in our earlier study (Kitai et al. 2007) using two unvaccinated horses that received the experimental infection with the same strain of WNV at an identical dose: One horse increased to ∼75% and another to 55%. Similarly, WNV-specific antibody responses were detected in vaccinated horses on days 9–21 (horse #5) or only on day 21 (horse #6). Horse #5 showed the maximum inhibition value of 49.5%, whereas horse #6 showed only 27.9%, which was close to the cutoff value. Although there were individual variations, these results indicated that the blocking ELISA can detect antibody responses to WNV in vaccinated horses. The WNV-specific antibody responses were detected in three (100%) and two (66.7%) of three horses in unvaccinated and vaccinated groups, respectively. The mean inhibition values were higher in unvaccinated than vaccinated horses (Fig. 2, left and middle panels). However, similar time courses of the mean inhibition values were obtained between three vaccinated and two unvaccinated horses, when horse #4 that did not develop detectable levels of NS1 antibody was excluded (Fig. 2, left and right panels). These results indicated no considerable effect of preimmunization with JE vaccine on WNV-specific NS1 antibody responses detected by the blocking ELISA after WNV infection, as far as NS1 antibody responses were induced in horses.
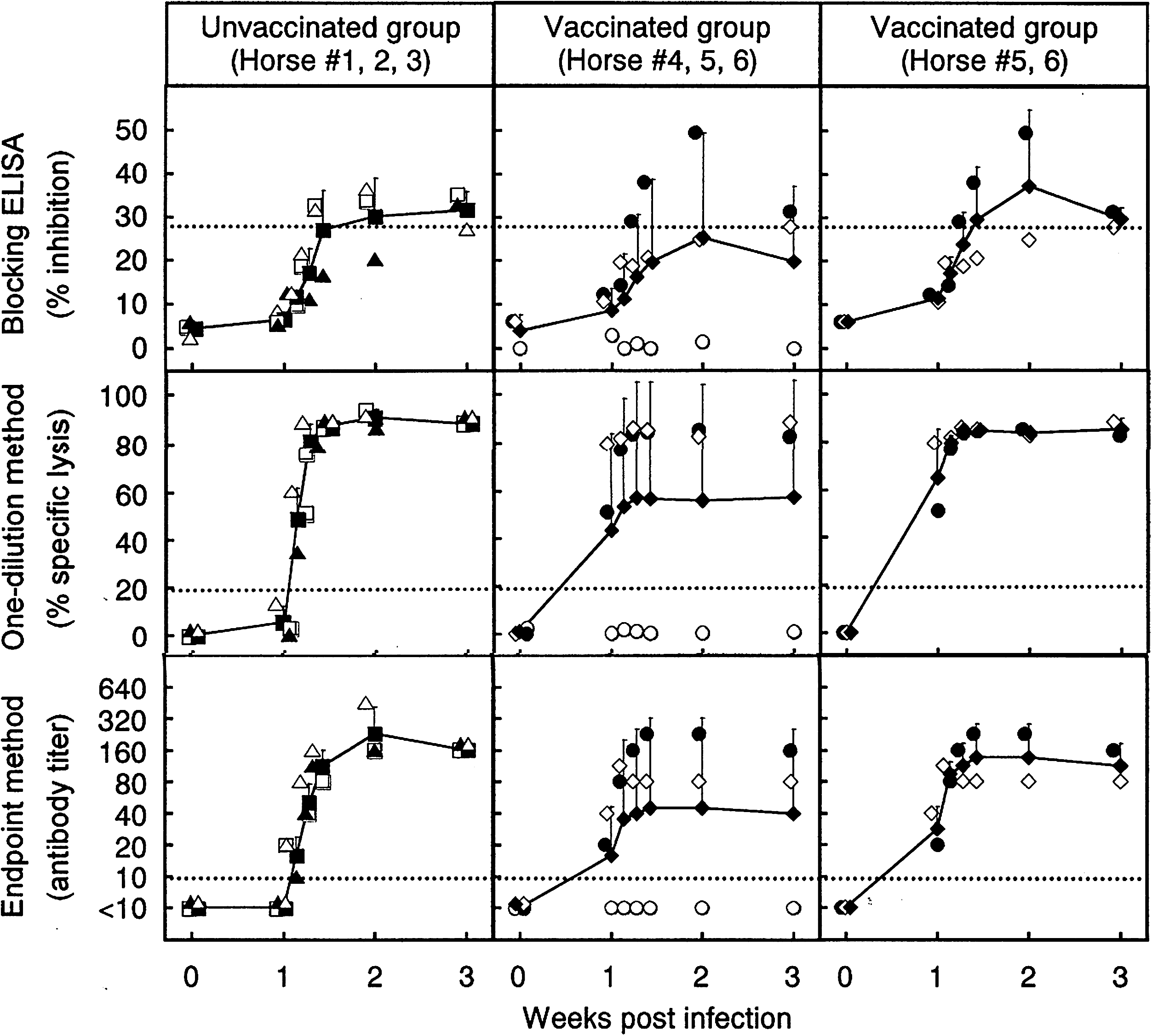
Comparison between the blocking ELISA and CDC assay using sera from WNV-infected horses with or without prevaccination with inactivated JE vaccine. Closed squares represent averaged data from three unvaccinated horses (horse #1, open triangle; horse #2, closed triangle; horse #3, open square), and closed diamonds represent averaged data from three or two vaccinated horses (horse #4, open circle; horse #5, closed circle; horse #6, open diamond). The data of individual horses are the same as those shown in Figure 1. Standard deviations of data are indicated by bars. Dotted lines indicate the cutoff value in the blocking ELISA (27.6%) and CDC assay (19.8% for the one-point method, 1:10 for the endpoint method).
CDC antibody levels
The effect of vaccination on NS1 antibody responses to WNV infection was examined by the CDC assay using both one-dilution and endpoint methods. The one-dilution method used percentages of specific lysis obtained with a 1:10 dilution of test sera (Fig. 1). In all unvaccinated horses, WNV-specific NS1 antibody responses were detected on day 8, and the percentages of specific lysis increased to >80% on day 10 and were maintained until the end of the experiment (day 21). In vaccinated horses, the percentages of specific lysis of horses #5 and 6 were already higher than the cutoff value (19.8%) on day 7 and were >80% on day 9 or after. Although data were not plotted, the percentages of specific lysis were <19.8% on day 6 in both horses #5 and 6, indicating that day 7 was the first day when CDC antibodies were detected. The antibody responses detected by the endpoint method were consistent with those of the one-dilution method in terms of the increasing pattern of antibody levels. The WNV-specific antibody responses were detected in three (100%) and two (66.7%) of three horses in unvaccinated and vaccinated groups, respectively. The time courses of the mean percentages of specific lysis/antibody titers obtained in unvaccinated horses were similar to those in vaccinated horses when the result from horse #4 was excluded from the calculation (Fig. 2, left and right panels). These results indicated that WNV-specific NS1 antibody responses could be detected by the CDC assay in both vaccinated and unvaccinated horses that induced antibody responses to WNV infection.
Discussion
In areas where JE is endemic, almost all horses are assumed to be immune to JEV. If WNV is introduced, this immunity may complicate the diagnosis of WNV infection. Horse sera used for the present blocking ELISA and CDC assay were previously tested by the neutralization test and MAC-ELISA (Shirafuji et al. 2009) (also see Fig. 1). Based on these data, similar antibody titers were shown against WNV and JEV in the neutralization test at most time points in horses vaccinated with the inactivated JE vaccine. The second exposure (to WNV antigen) induced strong anamnestic responses against the crossreactive antigens included in the first exposure (JE vaccine). A fourfold rise or higher in neutralizing antibody titer in paired sera collected in acute and convalescent phases is another criteria for diagnosis; this was observed against both WNV and JEV. Therefore, diagnosis will require both the neutralization test and MAC-ELISA against WNV and JEV: The presence of IgM antibody responses specific for WNV can demonstrate WNV infection. However, the levels of WNV-specific IgM antibody responses detected by MAC-ELISA (in which positive/negative ratios of ≥2.0 were regarded as positive) in vaccinated horses were lower than those in unvaccinated horses, shortening the period during which WNV-specific IgM antibody responses could be detected.
The abilities of the blocking ELISA (Kitai et al. 2007) and CDC assay (Kitai et al. 2010) to detect WNV-specific antibody responses have been already demonstrated using naïve horses. In the present study using vaccinated horses, comparison between the two new assays showed that the CDC assay was superior to the blocking ELISA for detecting WNV-specific NS1 antibody responses. The former provided high percentages of specific lysis in the one-dilution method or antibody titers in the endpoint method, whereas the latter provided only low inhibition values close to the cutoff value. The CDC assay detected WNV-specific antibody responses earlier than the blocking ELISA. In either assay, time courses were similar in vaccinated and unvaccinated horses, indicating that preimmunity to JE vaccine did not affect the ability of the assays to detect WNV-specific antibody responses, as far as NS1 antibodies were induced.
The better detection of WNV-specific NS1 antibody responses provided by the CDC assay may be attributed to the difference in assay variations between these methods. In the blocking ELISA, which is based on the binding inhibition of the marker antibody by a competitive antibody included in the test specimen, the variation in inhibition values obtained with control specimens that do not have WNV-specific antibodies is closely related to the cutoff value of the assay. Since four absorbance data are included in the formula to calculate the inhibition value [100−100 × (A−B)/(C−D): see Materials and Methods section for details], experimental variations appearing in ELISA seem to affect the calculation results. On the other hand, the CDC assay did not show so-called “non-specific” reactions in almost all negative controls, in our previous studies, to establish the assay method for detecting antibody responses to WNV (Kitai et al. 2010) and JEV (Konishi et al. 2008).
A prerequisite for an antibody assay for detecting infection is the induction of antibody responses after infection. In the present study, horse #4 that had been vaccinated and had a crossneutralizing antibody titer of 1:20 against WNV at the time of WNV infection did not show detectable NS1 antibody levels in the conventional and blocking ELISAs, as well as in the CDC assay, thus indicating that NS1 antigen was not sufficiently produced in this horse to induce NS1 antibody responses after inoculation with WNV. Since symptoms appeared on day 3 and neutralizing antibody titers increased (weakly but fourfold) on day 7 against JEV, or day 9 against WNV, the result does not seem to show “sterile” immunity, but to show that infection by the inoculated WNV was very limited in this horse. Crossprotection against WNV infection has been reported in JEV-immune mice (Lim et al. 2008), hamsters (Hammon and Sather 1956, Tesh et al. 2002), and monkeys (Goverdhan et al. 1992). The present study suggested that, to a certain degree, two doses of the inactivated JE vaccine protected horses from WNV infection with individual variations. Although only three horses were used for comparisons, fewer vaccinated horses developed clinical manifestations or RNAemia than unvaccinated ones, and the periods of clinical manifestations or RNAemia were shorter in vaccinated than unvaccinated horses. Thus, at least one limitation of the present new methods is the difficulty in detecting infection when crossimmunity reduces the induction of NS1 antibody responses; however, this would be the case for any antibody testing method.
In conclusion, the blocking ELISA, but preferably the CDC assay, appears useful for detecting WNV infection in horses and should support the present conventionally utilized methods. Although we used only vaccinated horses for the present evaluation, it is highly probable that these assays can specifically detect WNV infections also in horses that have a history of JEV infections, as horses including those naturally infected with JEV were used to obtain their cutoff values in our previous report (Kitai et al. 2007, 2010). These assays are rapid and do not need infectious agents and are, hence, advantageous over the neutralization test. Since only a small number of horses were used in the present study, further investigation will be needed to confirm the utility of these assays. The principle of these assays seems applicable to differential serodiagnosis between flaviviruses in areas where more than one flavivirus coexists.
Footnotes
Acknowledgments
This study was supported in part by grants from the Japan Racing Association, from Research on Emerging and Re-emerging Infectious Diseases, the Ministry of Health, Labour and Welfare of Japan, and from the Program of Founding Research Centers for Emerging and Reemerging Infectious Diseases, the Ministry of Education, Culture, Sports, Science, and Technology (MEXT), Japan.
Disclosure Statement
No competing financial interests exist.