
Abstract
Malaria accounts for the greatest morbidity and mortality of any arthropod-borne disease globally. Recently, it was determined that the protective antisporozoite CD8+ T-cell response originates predominantly from cutaneous lymph nodes draining the site of parasite inoculation by an Anopheles mosquito. The female mosquito inoculates sporozoites along with an assortment of salivary proteins into the skin of its mammalian host. Mosquito saliva has demonstrable antihemostatic as well as various immunomodulatory activities, and studies with mosquito-borne viruses support a role for mosquito saliva in enhancement of transmission and exacerbation of disease. Early differences in immune response can be detected, which discriminate between mice that are resistant and susceptible to neurological pathology. This supports the idea that early divergence in the immune response may influence the likelihood of progression to the more severe forms of malaria. To evaluate the effect of mosquito feeding on the pathogenesis and immune response to malaria, we injected washed Plasmodium berghei sporozoites intradermally in the presence or absence of mosquito feeding. We observed that mice exposed to mosquito feeding in tandem with the inoculation of sporozoites had higher parasitemias and an elevated progression to cerebral malaria. This was associated with, in particular, elevated levels of interleukin-4 and interleukin-10, suppression of overall transcription in response to infection, and decreased extravasation of dendritic cells and monocytes. This study enhances to our understanding of the complexity of the interactions between the malaria parasite, its host, and the mosquito vector.
Introduction
Most investigations of CM pathogenesis in murine models have primarily examined the immune processes relatively late in an infection initiated by inoculation of parasitized erythrocytes. In nature, exposure to malaria infection occurs via the bite of a Plasmodium-infected Anopheles mosquito, which inoculates the sporozoite stage of the parasite predominantly into intradermal and subcutaneous spaces (Amino et al. 2006a). Limited studies that utilizing intraperitoneal routes of infection suggest that early differences that discriminate between CM resistant and susceptible mice can be detected (Mitchell et al. 2005), supporting the notion that early divergence in the immune response may elevate or diminish the likelihood of progressing to the more severe forms of malaria. Importantly, mosquito saliva has demonstrated antihemostatic and immunomodulatory activities (Schneider et al. 2004, Wanasen et al. 2004, Ribeiro et al. 2007), and studies with mosquito-borne viruses support a role for mosquito saliva in enhancement of transmission and/or exacerbation of disease (Osorio et al. 1996, Edwards et al. 1998, Limesand et al. 2000, 2003, Schneider et al. 2006, Styer et al. 2006, Schneider and Higgs 2008). Preliminary studies in our laboratory have shown that after noninfectious mosquito bites, the saliva of Anopheles stephensi induces a rapid degranulation of cutaneous mast cells, an influx of neutrophils into the site of feeding, and lymph node hyperplasia (Demeure et al. 2005). Further, this Anopheles saliva-induced inflammatory response leads to a downregulation of subsequent T-cell-mediated immune responses mediated by interleukin (IL)-10 (Depinay et al. 2006).
Given the effect of the mosquito vector and the recent discovery that the protective antisporozoite CD8+ T-cell response originates predominantly from cutaneous lymph nodes (Chakravarty et al. 2007), assessing the immune response to mosquito-transmitted Plasmodium infection is of paramount importance, particularly at the site of inoculation and draining lymph nodes. Accordingly, we examined early immune responses to sporozoite inoculation and assessed the influence of mosquito saliva in this response and subsequent disease.
Materials and Methods
Mice
Naïve female C57BL/6 and BALB/c mice 5–6 weeks old were purchased from Charles River Breeding Laboratories (Saint-Aubin les Elbeufs, France). Mice were divided into three main groups: control mice inoculated with phosphate-buffered saline (PBS), mice inoculated with sporozoites alone, and mice inoculated with sporozoites and mosquito saliva, where mosquito saliva was either provided by injection of salivary gland extract (SGE) or feeding mosquitoes at the site of inoculation of sporozoites. No less than five mice were utilized per group and/or per time point. To evaluate whether the inoculation of saliva by mosquitoes causes detectable differences in the pathogenesis of murine malaria infection, C57Bl/6 mice were inoculated with 104 Plasmodium berghei sporozoites in the presence or absence of the feeding of ∼10 An. stephensi mosquitoes at the injection site (as determined by the presence of a blood meal) immediately before inoculation. Mosquitoes were starved for 24 h before blood feeding. This level of sporozoites was selected for consistently causing about 50%–70% CM. Inoculation (sporozoites or PBS control) and feeding were performed on a single footpad of each mouse. Mice were bled daily beginning on day 3 postinoculation via tail snip. A 4 μL sample of whole blood was added to 400 μL of PBS, and flow cytometry was utilized to detect the percent of parasitized red blood cells (out of 105 red blood cells) via fluorescence of the green fluorescent protein parasites. All animal care and experimentation were conducted in accordance with the Institut Pasteur animal care and use committee guidelines.
Mosquitoes and SGE
An. stephensi (sda 500 strain) mosquitoes were reared at the Center of Production and Infection of Anopheles (CEPIA) of the Institut Pasteur using standard procedures as reported earlier (Demeure et al. 2005). To obtain SGE, salivary glands were dissected from female An. stephensi mosquitoes as previously described (Schneider et al. 2004). When coinoculation was conducted, SGE was mixed with appropriate volumes of sporozoites, which were also maintained in a chilled PBS after isolation.
Parasites and mosquito infection
For all infections, P. berghei ANKA strain (Pb ANKA) lines expressing the green fluorescent protein on hsp70 promoter (Ishino et al. 2006) were used, allowing the detection of parasites throughout their life cycle in living mosquitoes and mice. This parasite induces experimental CM, characterized by paralysis, ataxia, convulsions, and coma between 7 and 12 days postinfection. Our prior studies with Pb ANKA demonstrate that mice succumb either between days 7 and 12 postinfection to CM or after day 20 postinfection to hyperparasitemia and anemia (Beghdadi et al. 2008, 2009). We used standard protocols for infecting and maintaining mosquitoes (Kebaier et al. 2010), which were infected by feeding upon gametocyte-carrying 6- to 8-week-old mice. Sporozoites used for transmission studies were isolated from mosquitoes 18 days after the infective blood meal. Before use of infected mosquitoes intravital fluorescence microscopy was employed on live, intact mosquitoes to establish that they had salivary gland infections; mosquitoes found to be negative were discarded.
Isolation of Pb ANKA sporozoites
Sporozoites were prepared by dissecting out and triturating infected salivary glands collected in sterile PBS, and washed twice by centrifugation at 12,000 g for 10 min to reduce the salivary gland component; previous studies have confirmed that such washing steps eliminated all detectable traces of apyrase and presumably mosquito saliva (Kebaier et al. 2010). Viability and concentration of sporozoites was determined microscopically. Sporozoites were maintained on ice and inoculated within 2 h of isolation.
RNA isolation and cytokine, chemokine, and sporozoite quantification in tissues
RNA was isolated from murine tissues as previously described (Schneider et al. 2004). To evaluate shifts of immune mediators and migration of sporozoites in tissues, C57BL/6 mice were exposed or not to mosquito feeding immediately before infection, and then at specific time points mice (five mice per group/replicate) were euthanized for sample collection. Tissues from uninfected mice were also processed as controls. Real-time RT-PCR utilized various primer-probe sets (Schneider et al. 2004) and followed standard TaqMan protocols (Applied Biosystems, Foster City, CA) (Schneider et al. 2004). Cytokine and chemokine expression was analyzed by the real-time RT-PCR at various early time points postinfection. Cytokine mRNA levels were normalized to hypoxanthine guanine phosphoribosyltransferase (HPRT) housekeeping gene expression. At two time points, 6 h in the skin and 24 h, in the draining lymph node expression of a large number of immune response-associated genes was assessed by the Th1-Th2-Th3 RT2 Profiler PCR Array (SABiosciences, Frederick, MD) according to the manufacturer's protocol.
To determine the rate of sporozoite migration, tissue samples were isolated at 1.5, 3, 6, and 24 h postinoculation of 104 sporozoites. For detection of sporozoites, RNA was isolated and expression of P. berghei 18S ribosomal RNA was quantified, normalized to HPRT gene expression, and related to known quantities of sporozoites (Prudencio et al. 2008). The real-time RT-PCRs to quantify P. berghei utilized Applied Biosystems Power SYBR Green PCR Master Mix and were performed according to the manufacturer's instructions on an ABI Prism 7000 system (Applied Biosystems).
Characterization of cell populations and migration in the skin and draining lymph node
Mice were divided into four groups based on the inoculum administered in the pinna of the ear: mice inoculated with PBS alone, 104 sporozoites alone, 104 sporozoites plus SGE, and 104 sporozoites after the feeding of ∼10 uninfected mosquitoes. Twenty-four hours after infection, ears and draining auricular lymph node were surgically removed. Tissues were reduced to single cell suspensions as previously described (Schneider et al. 2007). Blocking, staining, and cell characterization via multiparameter FACS analysis was performed as outlined previously (Beghdadi et al. 2008).
Statistical analysis
Statistical significance between samples was determined by Mann–Whitney test (GraphPad Prism; GraphPad Software, Inc., La Jolla, CA); values of p < 0.05 were considered significant. Significant differences in survival were evaluated by generation of Kaplan–Meier plots and log rank analysis (GraphPad Prism). p-Values of <0.05 were considered statistically significant.
Results
Elevated early parasitemia and increased progression to CM after coinoculation of sporozoites and mosquito saliva
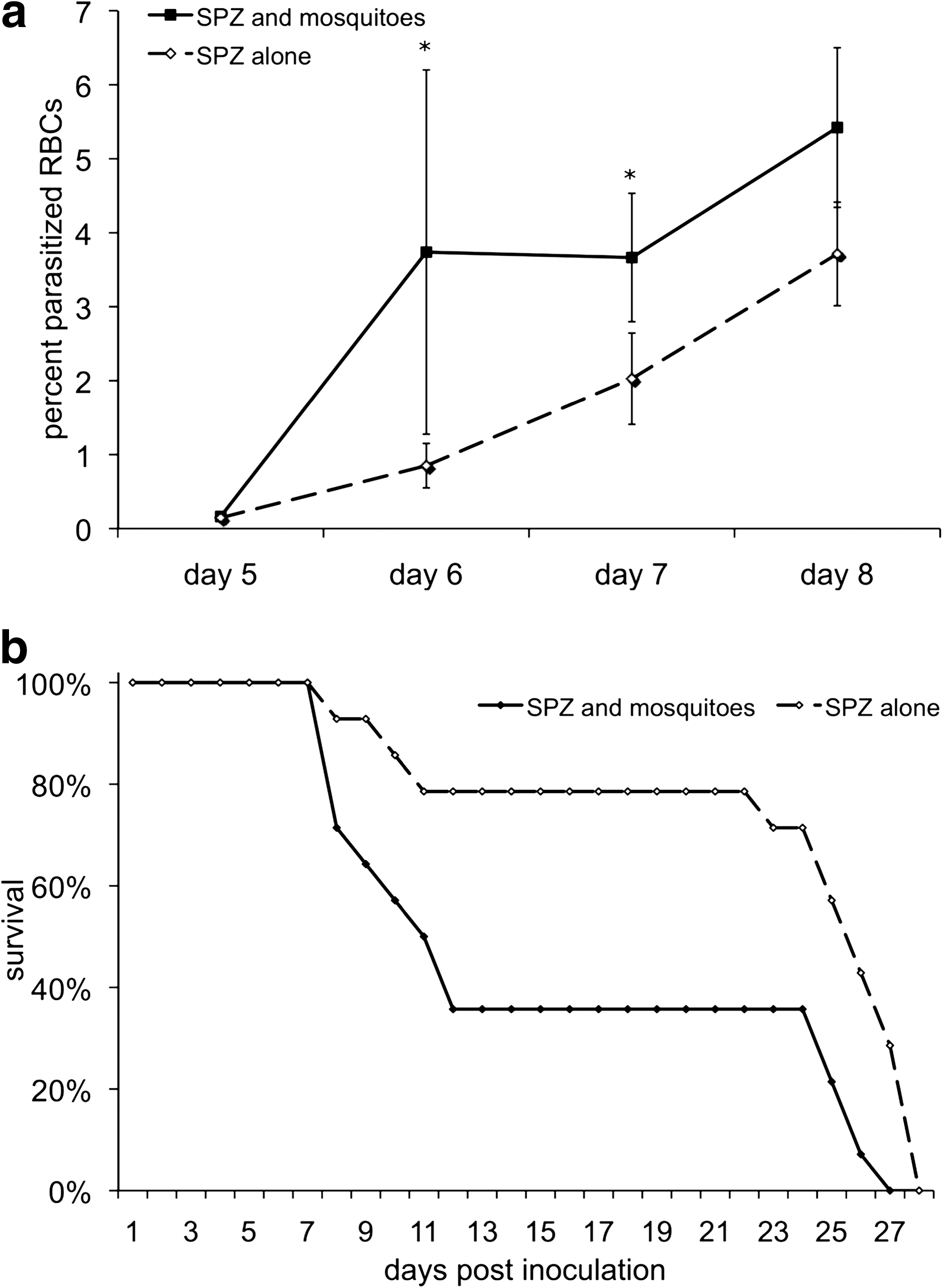
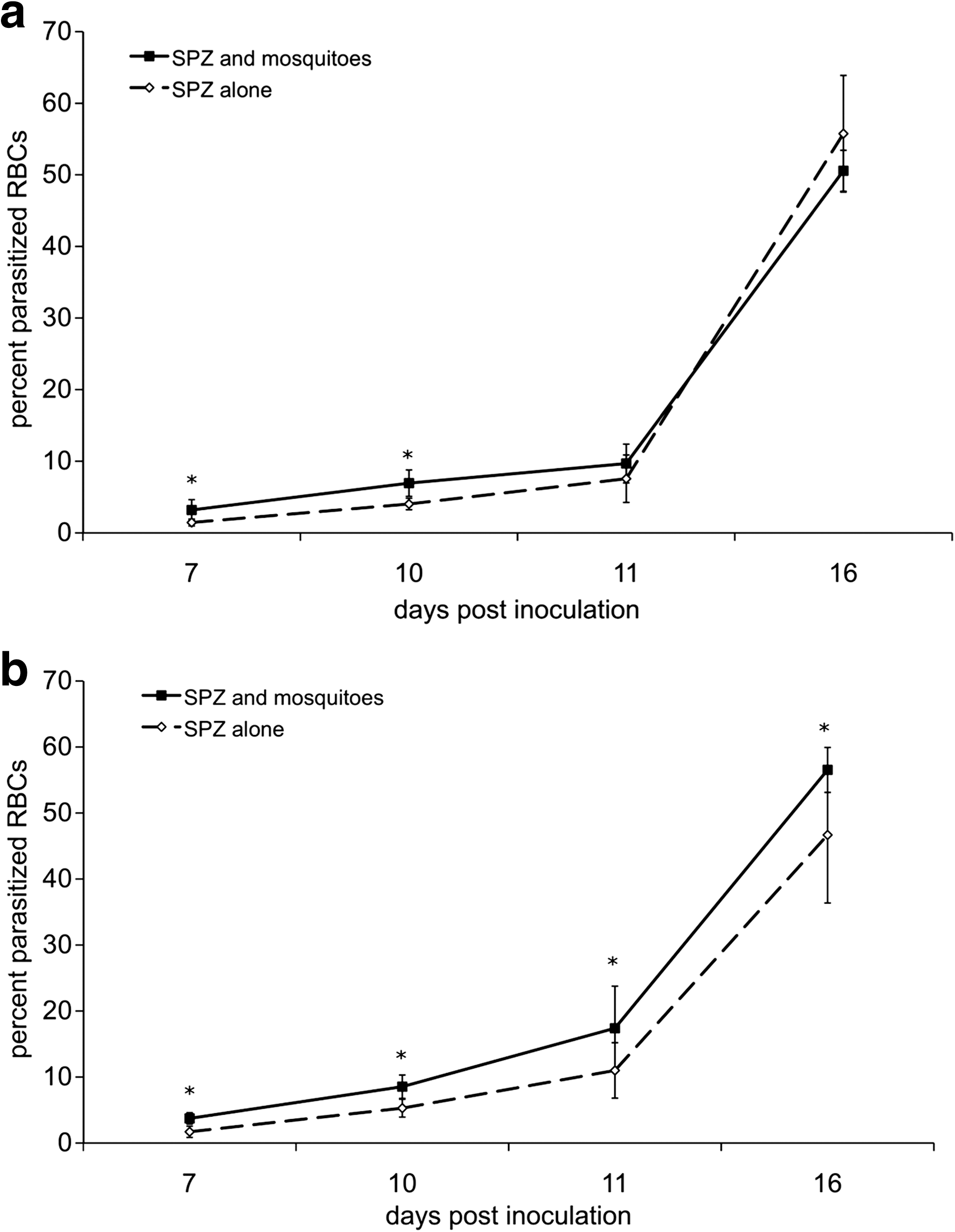
To evaluate whether the inoculation of saliva by mosquitoes causes detectable differences in the pathogenesis of murine malaria infection, mice were infected in the presence or absence of An. stephensi mosquito feeding at the injection site. We observed that mice exposed to mosquito feeding had higher parasitemias, particularly at early time points (Fig. 1a), when mice exposed to mosquitoes had nearly 4 times as many parasitized red blood cells. To ensure that the observed result was applicable to other mouse strains, a parallel experiment was also completed in BALB/c mice, which are resistant to infection and typically do not develop CM. BALB/c mice infected in the presence of mosquito feeding developed significantly higher parasitemias at either relatively low (1.5 × 104) or high (5.0 × 104) doses of P. berghei (Fig. 2a and b, respectively). Although the difference in parasitemias was statistically significant, the magnitude of the difference was relatively minor. BALB/c mice inoculated with <1.5 × 104 sporozoites generally did not progress to a detectable parasitemia.
Parasitemia
Parasitemia in BALB/c mice after intradermal inoculation of 1.5 × 104
After infection, mice were observed daily for signs of illness and vital status. Mice began to display recognizable signs of CM on day 8 and those that succumbed to CM normally died between days 9 and 12. Progressive signs of CM included ruffled fur, lethargy, hind limb weakness, ataxia, deviation of the head, convulsions, and coma. Significantly fewer mice injected with sporozoites alone progressed and succumbed to CM than mice infected after mosquito feeding (Fig. 1b and Table 1). It is important to discriminate between mice that succumbed to CM versus hyperparasitemis, as the mechanisms and effectors are not likely to be identical (Beghdadi et al. 2008). In all replicates of the experiment, mice that did not die from CM eventually succumbed to hyperparasitemia and anemia between days 20 and 27, with the exception of 2 mice from the group that were inoculated with sporozoites alone.
p-Values <0.05 were considered statistically significant.
Mice displaying characteristics signs of CM before death.
Significant differences in survival were evaluated by generation of Kaplan–Meier plots and log rank analysis.
CM, cerebral malaria; SPZ: Plasmodium berghei sporozite.
Sporozoite migration subsequent to dermal inoculation
One possible mechanism accounting for the observed difference in disease course is saliva-induced alteration in the migration or tropism of sporozoites either via altered motility of the parasite or a modulation of the host's inflammatory response. Thus, we first sought to establish the kinetics of sporozoite exit from the inoculation site and migration to the draining lymph node and/or liver. By 1.5 h postinoculation the number of sporozoites in the skin had decreased (Fig. 3a) and at this time point sporozoites were already detectable in the draining lymph node and liver (Fig. 3b, c). The number of sporozoites in the skin continually declined after inoculation, whereas in the draining lymph node a slight spike in sporozoites was observed at 3 h postinjection (Fig. 3b), consistent with an earlier study looking at migration of Plasmodium yoelii (Yamauchi et al. 2007). This early increase was more pronounced in the group of mice inoculated with sporozoites after An. stephensi feeding, though the difference was not significant. Detection of sporozoites was achieved utilizing molecular means; thus, the reduction in sporozoite levels may constitute either their emigration or destruction. Within the liver the number of sporozoites remained comparatively unchanged until 24 h postinoculation when the levels began to expand, likely due, not to continued migration, but rather the beginnings of replication in the organ (Fig. 3c). Interestingly, by 24 h postinoculation the level of sporozoite infection in the liver tended to be higher in the group where sporozoites were inoculated in the presence of mosquito saliva, although this difference was not statistically significant. Nonetheless, these data suggest that mosquito saliva does not affect the rate of sporozoite movement from peripheral sites to the liver.

Kinetics with which sporozoites migrate through skin
Mosquito feeding at the site of sporozoite inoculation mediates a suppression of local transcription
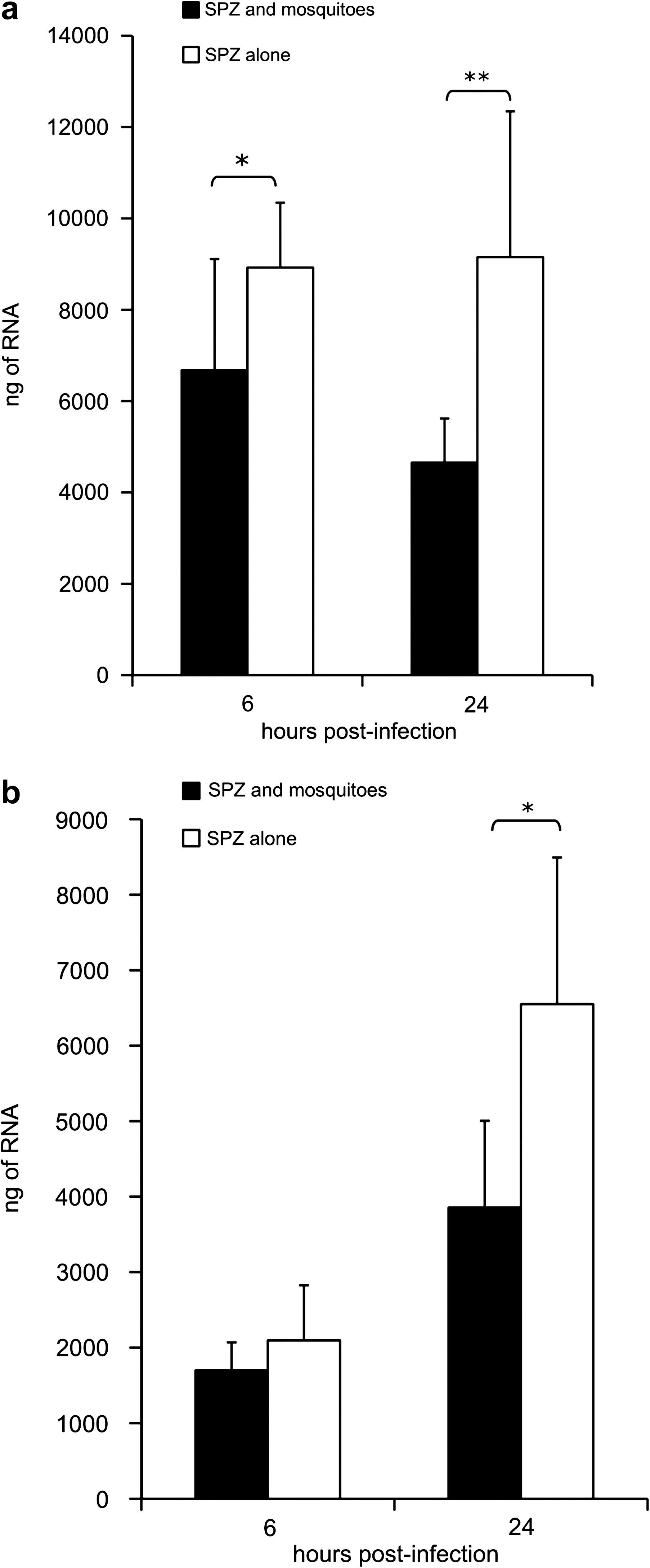
To investigate whether early disparity in tissue activation could help explain observed differences in P. berghei disease course, transcription as assessed by total RNA level was evaluated. The quantity of RNA was measured via spectrophotometry and normalized to the weight of the tissue sample. The total level (nanograms) of RNA was significantly less in the group infected after the feeding of mosquitoes (6 h: p < 0.05; 24 h: p < 0.01) (Fig. 4a). Expression of inoculation site RNA was nearly two times higher in the group injected with sporozoites alone (9.15 × 103 ± 3.19 × 103 as compared with 4.65 × 103 ± 0.97 × 103 for the group coexposed to mosquitoes and sporozoites; p < 0.01) by 24 h postinoculation. Additionally, the level of RNA expression in mice exposed to mosquitoes decreased, although not significantly, from 6 to 24 h by 30.3% (p = 0.06) whereas it remained stable or unchanged in mice inoculated with sporozoites alone. Differences in cell influx can only partially explain this observed disparity in RNA levels as suggested by the modest 16% increase in HPRT gene expression in mice inoculated with sporozoites alone (p = 0.1). Reduced mRNA expression could also be interpreted as an augmented efflux of skin resident cells or a pro-apoptotic effect elicited by mosquito bites. In draining lymph nodes there was no appreciable increase in RNA level at 6 h postinoculation, but by 24 h postinoculation both groups had significant amplification of RNA (sporozoites alone: p < 0.001; sporozoites and mosquito saliva: p < 0.005; Fig. 4b). Mice exposed to sporozoites alone had significantly higher RNA expression at 24 h postinfection than mice infected subsequent to mosquito feeding (p < 0.05). Overall, these results suggest a blunted response to infection in the skin and draining lymph nodes of mice exposed to mosquito feeding.
Host transcription activity as assessed by total RNA level. Skin
Mosquito saliva modulates cytokine expression during early P. berghei infection
A plausible cause of the shift on RNA levels in the skin and draining lymph nodes of mice exposed to mosquito feeding immediately before infection is modulation of immune signaling mediated by the pharmacologically active factors in mosquito saliva (Schneider and Higgs 2008). To explore this possibility we quantified expression levels of numerous cytokines and chemokines involved in immune cell activation, in the TH1- and TH2-type immune responses, in the antimicrobial humoral and other immune response-related genes by real-time RT-PCR. Genes of interest were individually verified in follow-up experiments at 6, 24, and/or 48 h postinfection (Table 2). Of the measured cytokines, IL-4 had the most pronounced alteration in expression level between infection groups. At 6 h postinoculation the level of IL-4 mRNA was 7.63 times higher (p < 0.05) in the inoculation site of mice exposed to mosquito saliva subsequent to sporozoite injection as compared with the mice injected with sporozoites alone. In this same group and time point IL-4 expression was 12.2 times higher (p < 0.05) in the draining lymph node, and this trend continued to the latest time point sampled, 48 h postinfection. IL-10 expression was similarly augmented in both the skin, 2.21 times at 6 h (p < 0.001), and draining lymph node, 1.96 (p < 0.05) and 1.49 times (p < 0.05) at 6 and 24 h postinfection, respectively (Table 2). At 6 h postinfection, a significant reduction in IL-3 expression was observed in the skin of the group that was exposed to both mosquito feeding and sporozoite inoculation as compared with the group injected with sporozoites alone (Table 2); inoculation site expression of IL-3 in mice exposed to mosquitoes was ∼1/3 of the level observed in mice injected with sporozoites alone (p < 0.05). At 24 h postinfection the draining lymph node of mice inoculated with sporozoites and mosquitoes had 2.2 times the level of IL-3 mRNA (p < 0.05) as the mice exposed to sporozoites alone (Table 2). Both cytotoxic T-lymphocyte-associated protein 4 (Ctla4) and IL-18 binding protein (IL-18bp) were significantly increased in the skin at 6 h postinfection by 30%–40% (p < 0.05; Table 2). Among the panel of genes analyzed, the most striking effect of mosquito bites was exerted on IL-4 and IL-10, suggesting a selective immune response in favor of parasite survival.
Values represent fold difference: SPZ and mosquitoes/SPZ alone in indicated tissues at specified times.
CD40, CD40 protein; CD69, CD69 protein; Ctla4, cytotoxic T-lymphocyte-associated protein 4; IFNb, interferon beta; IFNg, interferon gamma; IL-10, interleukin 10; IL-12, interleukin 12; IL-18bp, interleukin 18 binding protein; IL-1b, interleukin 1 beta; IL-3, interleukin 3; IL-4, interleukin 4; iNOS, inducible nitric oxide synthase; ND, not determined; Sftpd, surfactant associated protein D; Stat1, signal transducer and activator of transcription 1; Tbx21, T-box 21.
Indicated values significantly differed (p<0.05) between groups: SPZ and mosquitoes/SPZ alone.
Indicated values significantly differed (p<0.001) between groups: SPZ and mosquitoes/SPZ alone.
Altered inflammatory cell migration into early sites of infection in the presence of mosquito saliva
Given the known effects that vector saliva can have on cell migration (Ribeiro and Francischetti 2003, Schneider and Higgs 2008) and to further understand the decrease in total RNA expression, we characterized cell influx into the early peripheral sites of the antisporozoite immune response. Skin and draining lymph node populations were identified and their activation state was assessed. Whether exposed via mosquito feeding or coinoculation of SGE, sporozoite-infected, mosquito saliva-exposed mice had reduced recruitment of dendritic cells (by 54.2% and 57.7%, respectively; p < 0.05) 24 h postinfection at the inoculation site as compared with mice injected with sporozoites alone (Table 3). To a lesser degree, there was a tendency to have fewer monocytes in mice infected in the presence of mosquito saliva, with a reduction of 98.1% and 65.0% for mice exposed to sporozoites plus mosquitoes and sporozoites plus SGE, respectively, as compared with mice exposed to sporozoites alone (not statistically significant). A similar reduction in monocytes was observed at the same time point in the draining lymph node (12.8 × 104 ± 5.52 × 104 monocytes for sporozoite alone, vs. 7.97 × 104 ± 5.54 × 104 and 3.57 × 104 ± 1.09 × 104 for mice coexposed to mosquitoes or SGE, respectively), although only the group coinoculated with SGE was statistically different from the group injected with sporozites (SPZ) alone (p < 0.05). Additionally, eosinophils were more prevalent (approximately two to four times) within the skin of mice that had been exposed to mosquito saliva via either mechanism (p < 0.05). Overall, the effects of mosquito saliva on the cellular dynamics in the skin as compared with the inoculation of SPZ alone are a reduced recruitment of monocytes and dendritic cells, and an increased influx of granulocytes and eosinophils.
Mice were injected with 10,000 sporozoites alone, with 1.0 salivary gland pair equivalent of SGE, or immediately after the feeding of mosquitoes. The ears and draining lymph nodes were processed 24 h after inoculation, the cells obtained from both ears of each mouse were pooled, and the different populations of leukocytes were identified by staining and flow cytometry as described in the Materials and Methods section. The data shown are the mean ± 1 SD of five mice/group, and are representative of two independent replicates.
Values significantly different (p < 0.05) from the values of the group inoculated with SPZ alone. Values multiplied by 104 are the actual number of cells in each ear/lymph node.
SGE, salivary gland extract.
Discussion
This study demonstrates that An. stephensi feeding, and likely the inoculation of mosquito saliva specifically, potentiates Plasmodium infection in a murine model of malaria and, more widely, suggests that mosquito factors contribute to the pathogenesis of naturally transmitted malarial disease. Two distinct mouse models of malaria (C57BL/6 and BALB/c) displayed significantly elevated parasitemia after exposure to mosquito feeding, and, for the susceptible species, a higher progression to CM and associated mortality was observed. The present data suggest that mosquito feeding mediates these effects not by altering the tropism or migration of preerythrocytic parasites, but via dysregulation of immune signaling and a reduction in the recruitment of key inflammatory cells into the inoculation site. The significant length of time that sporozoites spend in the skin and draining lymph nodes, as demonstrated here and in other studies (Amino et al. 2006b, Kebaier et al. 2009), suggests that sporozoites are vulnerable to early inflammatory and immune responses in these tissues.
Mice exposed to An. stephensi feeding had higher parasite burdens and were more likely to succumb to CM. By utilizing the same inoculum for both groups and varying exposure to mosquitoes, the effect of mosquito feeding could be specifically addressed. Supplementary experiments suggest that the effect of mosquito feeding is most pronounced when it occurs directly before or after injection of parasites, and additional daily exposures to mosquitoes does not further exacerbate Plasmodium infection (data not shown). This adds support for the hypothesis that the effect of mosquito saliva is principally in the early sites of the infection, including the skin and draining lymph nodes. Nonetheless, the effect of mosquito feeding at a site distal to the sporozoite inoculation did not entirely abrogate its effect, suggesting that some of the effects of mosquito saliva are indeed systemic. This idea is supported by previous studies, including one that demonstrated an effect of mosquito feeding on splenocyte responses that persisted for up to 10 days postexposure (Zeidner et al. 1999).
These results clarify questions implied by a previous study (Vaughan et al. 1999) that found that sporozoites were more infectious to immunized mice when delivered by mosquito bite than by intravenous inoculation and estimated that one to two orders of magnitude fewer sporozoites were required to cause infection when delivered by mosquito, although this interpretation relies on assumptions on the numbers of sporozoites injected by mosquitoes. Our data suggest a plausible mechanism for enhanced infectiousness of sporozoites inoculated in the presence of mosquito saliva, namely, the early modulation of immune signaling and inflammatory cell extravasation. A recent study (Kebaier et al. 2010) demonstrated that the addition of SGE had no effect on sporozoite infectivity or the prepatent period; our results likewise observed a lack of difference in either of these indicators.
The present study suggests that the effect of mosquito saliva is not mediated via altered migration and, thus, higher primary infection of the liver, but rather the effect may be to perturb the early antiparasite immune response. The plausibility of this proposition is strengthened by a recent study that demonstrates that dendritic cells in cutaneous lymph nodes prime the first cohort of T cells after an infectious mosquito bite, and ablation of these lymphoid sites greatly impairs subsequent development of protective immunity against malaria (Chakravarty et al. 2007). This suggests that an alteration of the early immune response in the periphery, such as that observed in the present study, could effect downstream disease development. The most dramatic observed effect of mosquito saliva on immune signaling in this model was its ability to stimulate IL-4 expression both in vitro and in vivo. This concurs with previous studies on the effect of the saliva of other mosquito species (Zeidner et al. 1999, Schneider et al. 2004, 2006, Wanasen et al. 2004, Wasserman et al. 2004, Schneider and Higgs 2008). Recently, a novel mosquito salivary protein, SAAG-4, was shown to program host CD4+ T cells to express IL-4 while simultaneously reducing expression of interferon (IFN)-γ (Boppana et al. 2009). With many pathogens early cytokine responses involving IL-4 and IL-10 increase host susceptibility, whereas responses involving IL-12 and IFN-γ are important for resistance. Later in infection these roles are often reversed, as pro-inflammatory cytokines enhance immunopathology and regulatory cytokines mediate protection. Experiments with knock-out mice demonstrate that mice have higher resistance to P. berghei in the absence of IL-4 activity (Saeftel et al. 2004). Interestingly, this resistance to fatal disease could be abrogated if mice were infected with blood-stage or liver-stage parasites as apposed to infection via sporozoites, suggesting that the negative effect of IL-4 signaling is only in the very early stage of malaria infection and, thus, spatially and temporally linked with the mosquito-induced enhancement of IL-4 expression. Fonseca et al. (2007) similarly found a relative increase in IL-4 expression during Plasmodium chabaudi chabaudi infection after mosquito inoculation as compared with injection of blood stage parasites. Their study suggests that route of inoculation, parasite life-stage, or dose could influence this TH1 to TH2 shift. The present study allows the direct implication of mosquito feeding on this shift, by eliminating these variables and, additionally, suggests a role for enhanced IL-4 expression in the potentiation of malaria disease.
The upregulation of IL-10 expression after exposure to mosquito saliva as observed in this study agrees with previous studies in a range of mosquito species (Schneider et al. 2004, 2006, Depinay et al. 2006, Schneider and Higgs 2008). Although anti-inflammatory activity may be beneficial in later phases of malaria infection, during early infection expression of this cytokine may lead to a suppression of the TH1 response. In this regard, IL-10 inhibits the synthesis of multiple cytokines, including IFN-γ, IL-2, and TNF-β (Brady et al. 2003). In addition to antagonizing the function of IFN-γ and IL-12, IL-10 signaling downregulates major histocompatibility complex class II expression by monocytes and inhibits antigen presentation by several antigen-presenting cells (Enk et al. 1993, Macatonia et al. 1993). Even transient increases in IL-10 production can induce lasting T-cell inactivation that may limit the host's ability to control pathogens (Ejrnaes et al. 2006). In a murine model of malaria early IL-10 expression was associated with increased T regulatory cell proliferation, suppression of TH1 cytokines, as well as amplified parasitemia and mortality (Wu et al. 2007). Thus, the elevation of IL-10 expression by mosquito feeding may contribute to the worsened outcome of P. berghei infection in the presence of mosquito saliva.
The modulation of other signaling molecules also provides clues to the effect of mosquito feeding. Mice exposed to mosquito feeding concurrent with P. berghei infection demonstrated lower levels of IL-3 production in the skin at both 6 and 24 h postinfection, perhaps indicative of the suppression of inflammation in the inoculation site. Whereas, in the draining lymph node there was double the amount of IL-3 in mosquito-exposed mice. This increase in IL-3 may help to explain the increase in TH2 cytokines, as Ebner et al. (2002) demonstrated that dendritic cells cultured with IL-3 preferentially induced TH2 responses. Both Ctla4 and IL-18bp are significantly upregulated in the skin by mosquito feeding at 6 h postinfection. Ctla4 is a member of the immunoglobulin superfamily that is expressed on the surface of TH cells and transmits an inhibitory signal to T cells, and this protein is also found in regulatory T cells and may be important to their function (Noel et al. 1996). IL-18bp is an inhibitor of the proinflammatory cytokine IL-18. This protein binds to IL-18, prevents the binding of IL-18 to its receptor, and thus inhibits IL-18-induced IFN-γ production, resulting in reduced TH1 immune responses (Dinarello 2000). Despite the shifts in these signaling proteins, no significant alterations in IFN or IL-12 expression were detected.
The influx of key cells of the innate immune response in the site of sporozoite inoculation is significant in the early response to Plasmodium both for reducing the number of viable parasites that migrate to the liver and in initiating the adaptive immune response. A decrease in the recruitment of antigen presenting cells and inflammatory cells would thus be an advantage to the parasite, which is perhaps most vulnerable immediately after inoculation because of its low numbers and the drastic shift in environment to which it must adapt. The reduction in dendritic cells and lower trend in monocytes observed in the present study could hinder the early antiparasite response to P. berghei. Indeed, studies have previously demonstrated an important role for inflammatory cells in antiparasite activity (Waters et al. 1987, Pierrot et al. 2007). In addition to decreasing numbers of dendritic cells in the skin, it is possible that mosquito saliva alters phagocytosis of the sporozoites; in the present study we were unable to observe differences in the ability of mosquito saliva-treated peritoneal macrophages to internalize Fluoresbrite® yellow green carboxylate microspheres (Polysciences, Inc. Eppelheim, Germany; data not shown), although studies with ticks (Kyckova and Kopecky 2006) and sand flies (Zer et al. 2001) identified altered pathogen phagocytosis in the presence of saliva. In agreement with our study, an eosinophil chemotactic factor has recently been identified in anopheline mosquitoes (Owhashi et al. 2008). Presently, it is unclear whether eosinophil-mediated activity has a greater effect on control of malaria infection or the pathogenesis of severe malaria (Kurtzhals et al. 1998).
Collectively, this study extends previous research that demonstrates the important role that the mosquito can have on the transmission and pathogenesis of arthropod-borne agents (Schneider and Higgs 2008). The effect of mosquito saliva and mosquito transmission of pathogens has become recognized as an important factor in a diverse array of arthropod-borne infectious agents (Schneider and Higgs 2008). Much of this research has been conducted on arboviruses (Edwards et al. 1998, Limesand et al. 2000, Gillan and Devaney 2004, Schneider et al. 2006, Styer et al. 2006), although there is a large body of research on the role of sand fly saliva on Leishmania pathogenesis (Kamhawi 2000). In general these studies demonstrate that either previously resistant species become susceptible or susceptible species develop a more severe disease when mosquitoes or mosquito saliva are present during the introduction of the virus to the host, and often this is associated with a disregulation of the local immune response at the primary sites of viral replication (Limesand et al. 2003, Schneider et al. 2004, 2006, Wanasen et al. 2004, Wasserman et al. 2004, Depinay et al. 2006). It would stand to reason that the blocking of the effect of the mosquito salivary factors would thus provide protection against these agents by inhibiting the enhancing effect of mosquito saliva and/or by elevating the immune response at the site of inoculation. Whereas one study with P. yoelii showed that prior exposure to mosquitoes reduced early parasite development within the host (Donovan et al. 2007), we found that sensitizing mice to mosquito saliva before infection caused inconsistent results (data not shown); mice were either significantly protected, or disease was significantly exacerbated in mice pre-exposed to uninfected mosquitoes or SGE. We propose that the effect of sensitization to mosquito saliva is complicated by the activity of the many antigens in mosquito saliva as well as the immune response elicited by variations in prior exposure; in other words, an immune response to one mosquito protein may be advantageous to the host, whereas an immune response to another may benefit the pathogen. The plausibility of this has been verified by a recent study (Oliveira et al. 2008), which showed that immunization with different sand fly salivary proteins produced distinct immune profiles that correlated with resistance or susceptibility to Leishmania infection. Alternatively, the type of cellular response or the isotype of the antibody raised by sensitization to salivary antigens may prove important in dictating the consequence of prior exposure to mosquitoes.
In conclusion, our data demonstrate that An. stephensi feeding can potentiate disease in a recognized murine model of CM, suggesting that factors in mosquito saliva can play a role in malaria disease progression. The present study reveals that the exacerbation of Plasmodium infection is associated with elevated levels of IL-4 and IL-10, suppression of overall transcription in response to infection, and decreased extravasation of dendritic cells. Further research will add to our understanding of the complexity of the interactions between the malaria parasite, its host, and the mosquito vector.
Footnotes
Disclosure Statement
All authors declare that they have no commercial or other associations that might create a conflict of interest in connection with this article.