
Abstract
Choclo virus (CHOV) and Maporal virus (MAPV) are enzootic in Panama and western Venezuela, respectively. The results of previous studies suggested that the fulvous pygmy rice rat (Oligoryzomys fulvescens) is the principal host of both viruses. The results of an analysis of nucleotide sequence data in this study indicated that the rodent associated with CHOV is the Costa Rican pygmy rice rat (Oligoryzomys costaricensis) and that the rodent associated with MAPV is the delicate pygmy rice rat (Oligoryzomys delicatus). As such, MAPV is ecologically distinct from CHOV and should be considered a species separate from CHOV.
Introduction
The present-day principal host relationships of some hantaviruses appear to represent a long-term association between viruses in the genus Hantavirus and rodents in the family Cricetidae (Hjelle et al. 1995, Morzunov et al. 1998). Evidence for this ancient relationship includes the association of phylogenetically closely related hantaviral species with phylogenetically closely related, allopatric rodent species. For example, Bayou virus with the marsh oryzomys (Oryzomys palustris) in the southeastern United States (Ksiazek et al. 1997), Catacamas virus with Coues' oryzomys (Oryzomys couesi) in Honduras (Milazzo et al. 2006), and Playa de Oro virus with the Mexican oryzomys (Oryzomys mexicanus) in western Mexico (Chu et al. 2008).
The results of previous studies suggested that the fulvous pygmy rice rat (Oligoryzomys fulvescens) is the principal host of CHOV in Panama (Vincent et al. 2000, Suzàn et al. 2006, Armién et al. 2009) and Maporal virus (MAPV) in western Venezuela (Fulhorst et al. 2004, Milazzo et al. 2010). Yet, independent analyses of nucleocapsid protein gene sequences and glycoprotein precursor gene sequences indicated that MAPV is phylogenetically more closely related to ANDV than to CHOV (Richter et al. 2010). Hypothetically, the association of CHOV with the pygmy rice rat in Panama or the association of MAPV with the pygmy rice rat in western Venezuela is a consequence of a host-jumping event that occurred after the last common ancestor of the Panamanian and Venezuelan forms of O. fulvescens. This hypothesis assumes that the pygmy rice rat associated with CHOV virus and the pygmy rice rat associated with MAPV are conspecific.
Members of different species in the genus Oligoryzomys can be extremely difficult to distinguish from each other based on external physical characteristics alone (Dickerman and Yates 1995, Myers et al. 1995, Miranda et al. 2009). We note that the species identities of the CHOV-infected pygmy rice rats and MAPV-infected pygmy rice rats in previous studies (Vincent et al. 2000, Fulhorst et al. 2004, Milazzo et al. 2010) were based only on morphological features (i.e., length of tail, length and color of pelage, and skull topology).
The results of recent studies that included pygmy rice rats (Oligoryzomys spp.) captured in Mexico, Central America, Brazil, and Venezuela suggested that multiple species-level lineages exist within what is now regarded as O. fulvescens (Miranda et al. 2009, Rogers et al. 2009). Thus, the pygmy rice rat species associated with CHOV may be different from the pygmy rice rat species associated with MAPV. The objective of this study was to define better the taxonomic and phylogenetic relationship between the pygmy rice rat associated with CHOV in Panama and the pygmy rice rat associated with MAPV in western Venezuela.
Materials and Methods
Rodents
Pygmy rice rat MSB96073 was captured in 2000 on the Azuero Peninsula in Panama. The CHOV strain 588 (GenBank accession nos. DQ285046 and DQ285047) was originally isolated from the spleen of this rodent. Pygmy rice rats FHV-3968, FHV-3977, FHV-4083, and FHV-4110 were captured in 1997 during studies on the ecology of hantaviruses native to western Venezuela (Milazzo et al. 2010). FHV-3968 and FHV-3977 were captured near Caño Hondo in the State of Cojedes, FHV-4083 and FHV-4110 were captured near Caño Delgadito in the State of Portuguesa, and the MAPV prototype strain HV 97021050 was isolated from the spleen of FHV-4083 (Fulhorst et al. 2004). The carcasses of FHV-3968, FHV-3977, FHV-4083, and FHV-4110 were deposited into the Museo de Ciencias Naturales, Universidad Nacional Experimental de Los Llanos Occidentales “Ezequiel Zamora” under museum nos. B0999, B0991, B0901, and B0978, respectively. Samples of fresh-frozen liver from these rodents were deposited into the Museum of Texas Tech University under nos. TK138247, TK138248, TK138080, and TK138249, respectively.
Genetic characterization of rodents
The nucleotide sequence of a 976-bp fragment of the mitochondrial cytochrome-b (Cytb) gene of MSB96073 was determined previously (GenBank accession no. EU192164). The nucleotide sequences of the complete Cytb genes (1143-bp) of FHV-3968, FHV-3977, FHV-4083, FHV-4110, pygmy rice rat AMNH257263 (State of Sucre, Venezuela), pygmy rice rat MVZ155316 (Costa Rica), and pygmy rice rats CM76892 and CM76908 (Suriname) were determined in this study (Appendix 1). Genomic DNA was isolated from 0.1 g of liver, using a phenol extraction protocol (Longmire et al. 1997). The complete Cytb gene was amplified by using Go-Taq DNA Polymerase (Promega Corp., Madison, WI) in conjunction with oligonucleotides L14115 and H15288 (Martin et al. 2000). Both strands of the amplicon were sequenced directly, using the Big Dye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Inc., Foster City, CA). The nucleotide sequences of the Cytb genes of FHV-3968, FHV-3977, FHV-4083, FHV-4110, AMNH257263, MVZ155316, CM76892, and CM76908 were deposited into the GenBank nucleotide sequence database under accession nos. GU393995, GU393994, DQ227457, GU393996, GU393997, EU258539, EU258546, and GU393993, respectively.
Data analysis
The analyses of Cytb gene sequences included 29 rodents previously assigned to the O. fulvescens group (Carleton and Musser 1989, 1995), 39 other pygmy rice rats (Oligoryzomys spp.), a minute spiny mouse (Neacomys minutus), a forest small rice rat (Microryzomys minutus), and a Peruvian rice rat (Oreoryzomys balneator) (Appendix 1). Phylogenetic relationships were estimated using (1) a Markov chain Monte Carlo method implemented in the computer program MrBayes 3.1.2 (Huelsenbeck and Ronquist 2001, Ronquist and Huelsenbeck 2003), and (2) a maximum likelihood method implemented in PhyML (Guindon and Gascuel 2003) (
Results
The results of the Bayesian analyses (Fig. 1) were essentially the same as the results of the maximum likelihood analysis (phylogram not shown, bootstrap values >70% shown in Fig. 1) and indicated that the pygmy rice rat associated with MAPV, the pygmy rice rat associated with CHOV, and the fulvous pygmy rice rat (O. fulvescens) represent different phylogenetic lineages in the genus Oligoryzomys. Pygmy rice rat FHV-4083 and the other pygmy rice rat from Portuguesa (i.e., FHV-4110) were included in a clade (labeled “A” in Fig. 1) that contained the pygmy rice rats from Cojedes, the pygmy rice rat from Sucre, and the delicate pygmy rice rat (Oligoryzomys fulvescens delicatus) from Suriname (i.e., CM76908). Monophyly of the delicate pygmy rice rat from Suriname and the five other members of clade A was supported by probability values and the results of the bootstrap analyses. Clade A was sister to Oligoryzomys messorius, monophyly of the three specimens of O. messorius was supported by probability values and the results of the bootstrap analyses, and monophyly of clade A and O. messorius was supported by probability values. Pygmy rice rat MSB96703 and the two other pygmy rice rats from Panama were included in a clade (labeled “B” in Fig. 1) that contained the Costa Rican pygmy rice rat (Oligoryzomys fulvescens costaricensis) from Costa Rica (i.e., MVZ155316). This clade was sister to a clade that contained Oligoryzomys vegetus and O. fulvescens. Monophyly of the three rodents in clade B and monophyly of (1) O. fulvescens, (2) O. vegetus, and (3) O. fulvescens and O. vegetus were supported by clade probability values calculated a posteriori and the results of the bootstrap analyses; however, monophyly of clade B, O. vegetus, and O. fulvescens was not supported by probability values or the results of the bootstrap analyses.
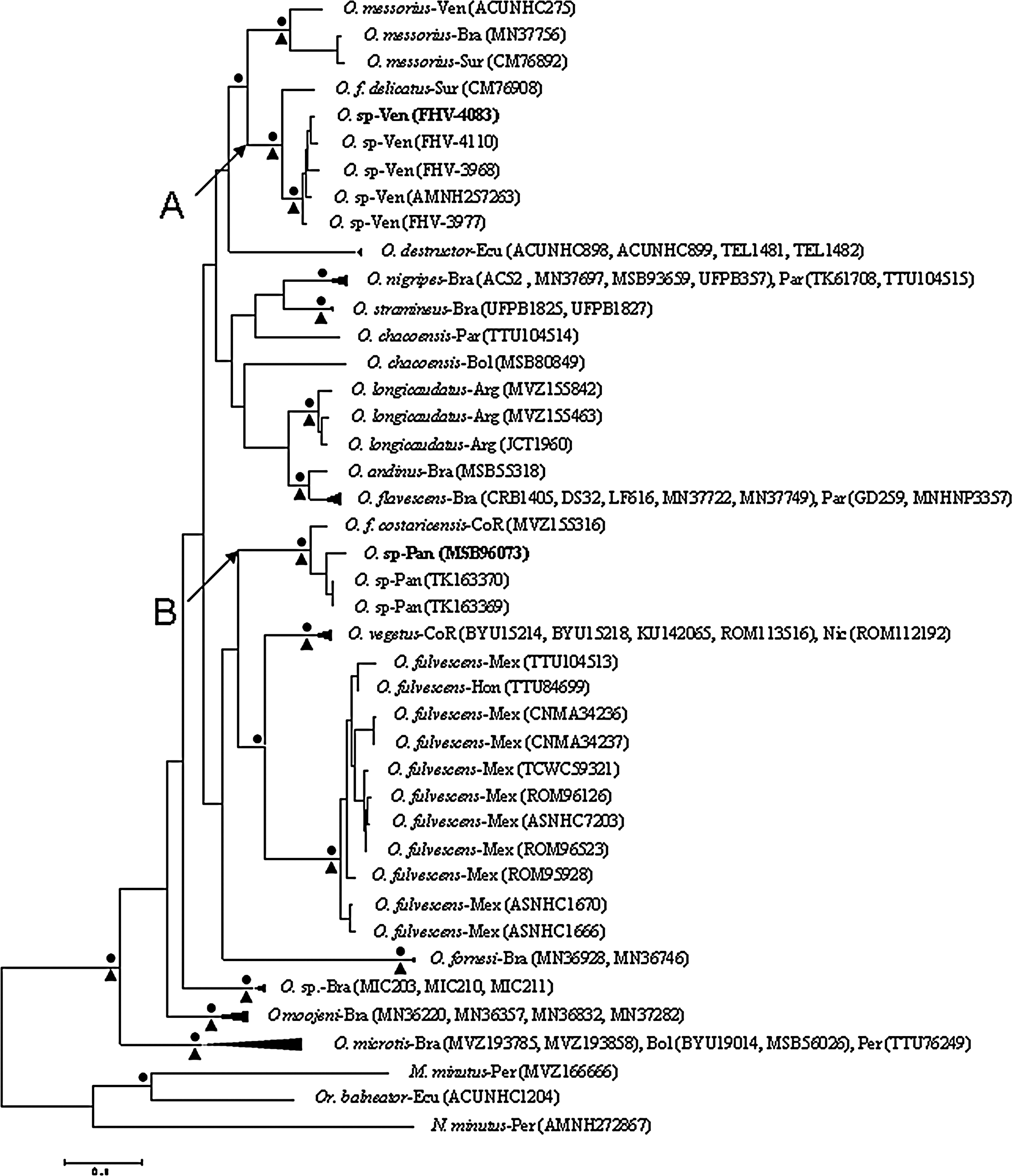
Phylogenetic relationships among rodents based on Bayesian analyses of mitochondrial cytochrome-b gene sequences. The results of the maximum likelihood analysis were essentially the same as the results of the Bayesian analyses. The scale bar indicates the expected number of substitutions per site. The black dots (•) at the nodes indicate clade probability values ≥95.0%. The black triangles (▴) indicate bootstrap support values ≥70%. Bootstrap values for clades A and B were 99% and 100%, respectively. Probability values and bootstrap support values are not listed for nodes below species level. Branch labels include species name–country (museum number). Arg, Argentina; Bol, Bolivia; Bra, Brazil; CoR, Costa Rica; Ecu, Ecuador; Hon, Honduras; Mex, Mexico; Nic, Nicaragua; Pan, Panama; Par, Paraguay; Per, Peru; Sur, Suriname; Ven, Venezuela. The Maporal virus–infected pygmy rice rat (FHV-4083) and Choclo virus–infected pygmy rice rat (MSB96073) are in boldface.
In pairwise comparisons of Cytb gene sequences, genetic distances between different Oligoryzomys spp. ranged from 4.45% to 12.09%. Further, the genetic distances between the pygmy rice rats from Portuguesa and the delicate pygmy rice rat from Suriname (CM76908) ranged from 3.70% to 3.98%, the genetic distance between MSB96703 and Costa Rican pygmy rice rat MVZ155316 was 3.13%, the genetic distances between the pygmy rice rats from Portuguesa and MSB96703 ranged from 9.96% to 10.07%, and the genetic distances between clade A and O. fulvescens and between clade B and O. fulvescens ranged from 7.97% to 10.57% and 9.81% to 11.88%, respectively (Table 1).
Mean (range) of the Kimura 2-parameter genetic distances (Kimura 1980), calculated using MEGA4 (Tamura et al. 2007).
Clade A includes the delicate pygmy rice rat (Oligoryzomys delicatus) from Suriname and the pygmy rice rats from Portuguesa (Venezuela). Clade B includes the Costa Rican pygmy rice rat (Oligoryzomys costaricensis) from Costa Rica and three pygmy rice rats from Panama (see Fig. 1).
Discussion
Collectively, the results of the Bayesian analyses, maximum likelihood analysis, and pairwise comparisons of Cytb gene sequences suggest that clades A and B represent different species within the genus Oligoryzomys and that these species are distinct from O. fulvescens. These results are similar to the results of a recently published study (Rogers et al. 2009). The oldest available name for the members of clade A is Oligoryzomys delicatus (Allen and Chapman 1897). Similarly, the oldest available name for members of clade B is Oligoryzomys costaricensis (Allen 1893). Pending a morphological review of the type specimens and formal taxonomic revision of the genus Oligoryzomys, the rodent associated with MAPV in western Venezuela should be referred to as the delicate pygmy rice rat (O. delicatus, alternatively—O. fulvescens delicatus) and the rodent that serves as the principal host of CHOV in Panama should be referred to as the Costa Rican pygmy rice rat (O. costaricensis, otherwise—O. fulvescens costaricensis).
The Eighth Report of the International Committee on Taxonomy of Viruses (Nichol et al. 2005) indicated that different hantaviral species should occupy different ecological niches. Together, the association of MAPV with O. delicatus and the association of CHOV with O. costaricensis indicate that MAPV and CHOV occupy different ecological niches. As such, the results of this study support the notion that MAPV is a species distinct from CHOV (Fulhorst et al. 2004).
The results of analyses of nucleotide sequence data in a recent study (Richter et al. 2010) indicated that MAPV is phylogenetically more closely related to ANDV than to CHOV. The results of the Bayesian and maximum likelihood analyses of Cytb gene sequences in this study suggest that O. delicatus is phylogenetically more closely related to O. longicaudatus (the principal host of ANDV) than to O. costaricensis. Although the close relationship between O. delicatus and O. longicaudatus was not statistically supported in this study, the association of MAPV with O. delicatus in western Venezuela together with the association of ANDV with O. longicaudatus in Argentina (Levis et al. 1998) and Chile (Medina et al. 2009) suggests that the association of MAPV with O. delicatus was established long ago.
Footnotes
Acknowledgments
We thank Duke S. Rogers (Brigham Young University, Provo, Utah) and Brenda Rodgers (Texas Tech University) for comments on drafts of this article. Samples of liver were obtained from Abilene Christian University Natural History Collections (Abilene, Texas), American Museum of Natural History (New York, New York), Louisiana State University Museum of Natural History (Baton Rouge, Louisiana), and The Museum of Texas Tech University (Lubbock, Texas). Funding was provided by the Department of Biology, Texas Tech University and National Institutes of Health (NIH) grant AI-067947. Laboratory space at Texas Tech University was provided by Brenda Rodgers.
Disclosure Statement
No competing financial interests exist.
Appendix 1. Rodents Included in the Analysis of Cytochrome- b Gene Sequences
| Species | Rodent (voucher specimen number, GenBank accession number) a |
|---|---|
| Oligoryzomys andinus | Bolivia—MSB55318 (NK12051, GU393999) |
| Oligoryzomys chacoensis | Bolivia—MSB80849 (NK72388, EU192173), Paraguay–TTU104514 (TK62932, EU258543) |
| Oligoryzomys costaricensis | Costa Rica—MVZ155316 (LSUMZ4745, EU258539 b ); Panama–MSB96073 (NK101588, EU192164), TK163369 (GU393988), TK163370 (GU393989) |
| Oligoryzomys delicatus | Suriname—CM76908 (TK21065, GU393993 b ); Venezuela—AMNH257263 (HGC773, GU393997 b ), FHV-3968 (TK138247, GU393995 b,c ), FHV-3977 (TK139248, GU393994 b,c ), FHV-4083 (TK138080, DQ227457 b,c ), FHV-4110 (TK138249, GU393996 b,c ) |
| Oligoryzomys destructor | Ecuador—ACUNHC898 (TEL1479, EU258544), ACUNHC899 (TEL1526, GU393992), TEL1481 (GU393991), TEL1482 (GU393990) |
| Oligoryzomys flavescens | Brazil—CRB1405 (EU258545), DS32 (DQ826006), LF616 (DQ826009), MN37749 (DQ826015), MN37722 (DQ826014); Paraguay—GD259 (EU192158), MNHNP3357 (TK64399, EU258542) |
| Oligoryzomys fornesi | Brazil—MN36746 (DQ826022), MN36928 (DQ826023) |
| Oligoryzomys fulvescens | Honduras—TTU84699 (TK102042, EU258547); Mexico—ASNHC1666 (EU294234), ASNHC1670 (EU294232), ASNHC7203 (EU294236), CNMA34236 (EU294248), CNMA34237 (EU294247), ROM95928 (EU294229), ROM96126 (EU294238), ROM96523 (EU294240), TCWC59321 (EU294242), TTU104513 (TK138057, EU258548) |
| Oligoryzomys longicaudatus | Argentina—JCT1960 (EU192160), MVZ154463 (GU393998), MVZ155842 (U03535) |
| Oligoryzomys messorius | Brazil—MN37756 (DQ826025), Suriname—CM76892 (TK17858, EU258546 b ), Venezuela—ACUNHC275 (EU258537) |
| Oligoryzomys microtis | Bolivia—BYU19014 (AY439000), MSB56026 (NK13425, EU192172); Brazil—MVZ193785 (MNFS1321, U58381), MVZ193858 (EU258549); Peru—TTU76249 (FJ374766) |
| Oligoryzomys moojeni | Brazil—MN36220 (DQ826016), MN36357 (DQ826017), MN36832 (DQ826019), MN37282 (DQ826020) |
| Oligoryzomys nigripes | Brazil—AC52 (DQ825987), MN37697 (DQ826003), MSB93659 (NK42266, EU192163), UFPB357 (DQ826004); Paraguay—TK61708 (EU258551), TTU104515 (TK65938, EU258550) |
| Oligoryzomys stramineus | Brazil—UFPB1825 (DQ826026), UFPB1827 (DQ826027) |
| Oligoryzomys vegetus | Costa Rica—BYU15214 (EU294250), BYU15218 (EU294249), KU142065 (EU192165), ROM113516 (EU258541); Nicaragua—ROM112192 (EU258538) |
| Oligoryzomys sp. | Brazil—MIC203 (EU192169), MIC210 (EU192167), MIC211 (EU192168) |
| Microryzomys minutus | Peru—MVZ166666 (EU258535) |
| Neacomys minutus | Peru—AMNH272867 (EU258536) |
| Oreoryzomys balneator | Ecuador—ACUNHC1204 (TEL1854, EU258534) |
Voucher specimen (tissue) number if different from the museum number assigned to the rodent.
Cytb gene sequence determined in this study.
The carcasses of FHV-3968, FHV-3977, FVH-4083, and FHV-4110 were deposited into Museo de Ciencias Naturales, Universidad Nacional Experimental de Los Llanos Occidentales “Ezequiel Zamora” under museum nos. B0999, B0991, B0901, and B0978, respectively.
ACUNHC, Abilene Christian University Natural History Collections (includes specimens identified with TEL); AMNH, American Museum of Natural History (includes the specimen identified with HGC); ASNHC, Angelo State Natural History Collections; BYU, Monte L. Bean Museum, Brigham Young University; CM, Carnegie Museum of Natural History; CNMA, Colección Nacional de Mamíferos, Universidad Nacional Autonóma de México; KU, University of Kansas Natural History Collection; LSUMZ, Louisiana State University Natural History Collection; MN, Museum Nacional do Rio de Janeiro (includes the specimens identified with AC, CRB, DS, GD, JCT, LF, MIC, or UFPB); MNHNP, Museo Nacional de Historia Natural de Paraguay; MSB, Museum of Southwestern Biology (includes the specimens identified with NK); MVZ, Museum of Vertebrate Zoology (includes the specimen identified with MNFS); ROM, Royal Ontario Museum; TCWC, Texas Cooperative Wildlife Collection; TTU, Museum of Texas Tech University (includes the specimens identified with TK). Museum acronyms follow Hafner et al. (1997).