
Abstract
Borrelia burgdorferi, the bacterium that causes Lyme disease, is transmitted to a susceptible host by Ixodes spp. tick bites. However, there is uncertainty whether B. burgdorferi are shed from ticks by the fecal route. In this study, B. burgdorferi–infected ticks were fed on mice while confined to a certain area of the skin by a capsule. During and after feeding, tick feces were collected and placed in Barbour-Stoenner-Kelley (BSK)-II media for cultivation and in sterile water for polymerase chain reaction (PCR) analysis. Although none of the tested samples were culture positive for B. burgdorferi, all but one of the fecal DNA samples from infected ticks were PCR positive. These results indicated that B. burgdorferi were shed from feeding ticks during defecation and suggest that the spirochetes did not remain viable once exposed to the outside environment. This finding has important ramifications for investigators interpreting B. burgdorferi–specific PCR results when conducting tick transmission experiments.
Introduction
In a separate study, we performed polymerase chain reaction (PCR) for detection of borrelial DNA within mouse skin at the tick bite site and observed a copious amount of tick feces upon the external feeding area. Therefore, to eliminate the consideration of any PCR positives being the result of fecal contamination, we conducted the current study to determine whether B. burgdorferi were present in the feces of feeding ticks.
Materials and Methods
Strains and ticks
The wild-type (WT) B. burgdorferi strain used was B31-A3, a clonal infectious strain (Elias et al. 2002). bba64::flgkan is a mutant strain of the B31-A3 parental strain whereby the bba64 gene was inactivated, and strain bba64::flgkan-ciscomp is the complemented mutant strain as described previously (Gilmore et al. 2010). B. burgdorferi were cultivated in Barbour-Stoenner-Kelley (BSK)-II (Barbour 1984) complete medium at 34°C in capped tubes. Ixodes scapularis ticks were infected separately with clonal strains of B. burgdorferi WT, bba64 mutant (bba64::flgkan), and bba64 complemented mutant (bba64::flgkan-ciscomp) by feeding clean larvae on infected mice as described previously (Piesman 1993, Gilmore et al. 2010). The newly infected larvae were allowed to molt to the nymphal stage (the Borrelia persisted transtadially), and were used for subsequent feedings on mice.
Tick feedings
Infected nymphal ticks were placed in plastic capsules affixed to 6–8-week-old female Swiss-Webster outbred mice (specific pathogen-free colony maintained at the Division of Vector-Borne Diseases, Fort Collins, CO) to feed within a localized area of the skin as follows. Mice were anesthetized and a small area on the dorsal surface between the shoulder blades was shaved. A feeding capsule (18 mm in diameter; Nalgene) was glued to the skin using a mixture of three parts colophony (Kramer Pigments Inc.) and one part beeswax. Nymphal ticks were placed into the capsule, and a small piece of mesh was attached to the top (Mbow et al. 1994). The mesh screen prevented tick escape from the contained area of skin and was removed to collect feces at specific times and to allow ticks to crawl out once fed to repletion. Uninfected adult tick feedings were conducted on female New Zealand white rabbits (Western Oregon Rabbit Co.) (Ledin et al. 2005). Approximately 50 female ticks were fed on the ears of each rabbit and were contained within cloth bags held in place by tape at the bottom of the ear. The mouse and rabbit experimental protocols were approved by the Division of Vector-Borne Diseases Animal Care and Use Committee.
Fecal collection for PCR and culture
The first set of fecal samples were collected individually from 11 mice following tick repletion, ∼4 days after tick attachment. One of the 11 mice was fed upon by WT-infected nymphs (n = 10), 1 mouse was fed upon by bba64 mutant-infected nymphs (n = 12), and 9 mice were fed upon by nymphs infected with the complemented mutant (10 ticks/mouse). The second set of fecal samples came from WT-infected nymphs (n = 11) and was collected at the capsule site from a single mouse each day of the feeding; that is, fecal samples were obtained at 30, 54, 76, and 96 h postinfestation.
Tick fecal samples were collected from within capsules by swabbing the area with a cotton-tipped swab dampened with sterile water or BSK medium. Each swab was then dispensed into a tube of sterile water (0.5 mL). Before placement in the water tube, 5 BSK-dampened swabs were first inoculated into BSK-II complete medium supplemented with antibiotics and Fungizone (Sinsky and Piesman 1989). Feces were collected from uninfected adult ticks after the ticks fed on rabbits. After tick repletion, dry feces were collected from the cloth bags used to contain the ticks that covered the rabbits' ears. A sampling of uninfected feces were suspended in water in preparation for PCR. Collection tubes containing fecal samples were stored at −20°C.
PCR and DNA sequencing
DNA was purified from 100 μL of the tick fecal water suspensions using the QIAamp DNA Micro Kit (Qiagen) and eluted into 30 μL final volume with water. PCR was performed using 5 μL of the purified DNA samples using RubyTaq PCR Master Mix (USB) and primers specific for the flaB (Johnson et al. 1992) and bba64 (Gilmore et al. 2010) genes of B. burgdorferi in a 20 μL total volume. Amplification parameters were 94°C for 1 min followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 60 s, and 1 cycle of 72°C for 5 min. Amplicons were observed by agarose gel electrophoresis loading 5 μL of the PCR reaction. Amplicons were purified from the PCR reaction mix using QIAquick PCR purification Kit (Qiagen), and were sequenced using Big Dye Terminator Cycle Sequencing Kit ver. 3.1 (Applied Biosystems).
Results
PCR analyses of tick fecal DNA
PCR was performed on DNA isolated from tick feces obtained from the 11 capsule feedings with WT-, bba64 mutant-, and complement-infected ticks using B. burgdorferi flaB primers. Amplicons were detected in 8/9 samples from the complement-infected ticks, and from both samples (2/2) from the WT- and mutant-infected ticks (10/11 total samples) (Table 1). No amplicons were detected from feces obtained from uninfected ticks. The PCR assays yielded qualitative results since the collections of fecal material were not a quantitative process. Failure to detect an amplicon in one of the complement-infected tick samples may have been due to a lack of sensitivity for that sample. Additionally, the DNA samples were tested for the bba64 gene by PCR, and all samples that were PCR positive for flaB were also positive for bba64 (Table 1). The flaB and bba64 amplicons were sequenced and demonstrated 100% identity with the respective strain B31 genes.
Feces collected from 8 to 12 ticks/mouse.
Tick feces pooled from feedings on rabbits.
PCR, polymerase chain reaction; nd, not done; WT, wild type.
The number of ticks placed into the mouse capsules to feed and the number of ticks recovered following repletion are recorded in Table 2. From the capsule experiments, we observed an average of 90% tick attachment, with the exception of two instances, whereby the capsule became detached and the majority of the ticks were groomed off. No >2 ticks/capsule were unaccounted for, with all ticks recovered from three mice.
Ticks not contained by a capsule can be groomed off and destroyed by the mouse.
We repeated the experiment using WT-infected ticks on a single mouse and collected fecal matter once a day during the feed instead of solely at tick repletion as was done in the first experiment. As a control, the area of mouse skin within the capsule was swabbed before placement of the ticks. After tick infestation, fecal samples were collected at 30, 54, 76, and 96 h. The flaB gene was detected by PCR from each time point sample (Fig. 1). Although reasonable to conclude that B. burgdorferi were shed throughout the feed, it was not possible to determine absolutely whether organisms were shed continuously because the first time point sample at 30 h was PCR positive. In other words, the later samples were positive due to either continous borrelial shedding, and/or from DNA present in the early sample.
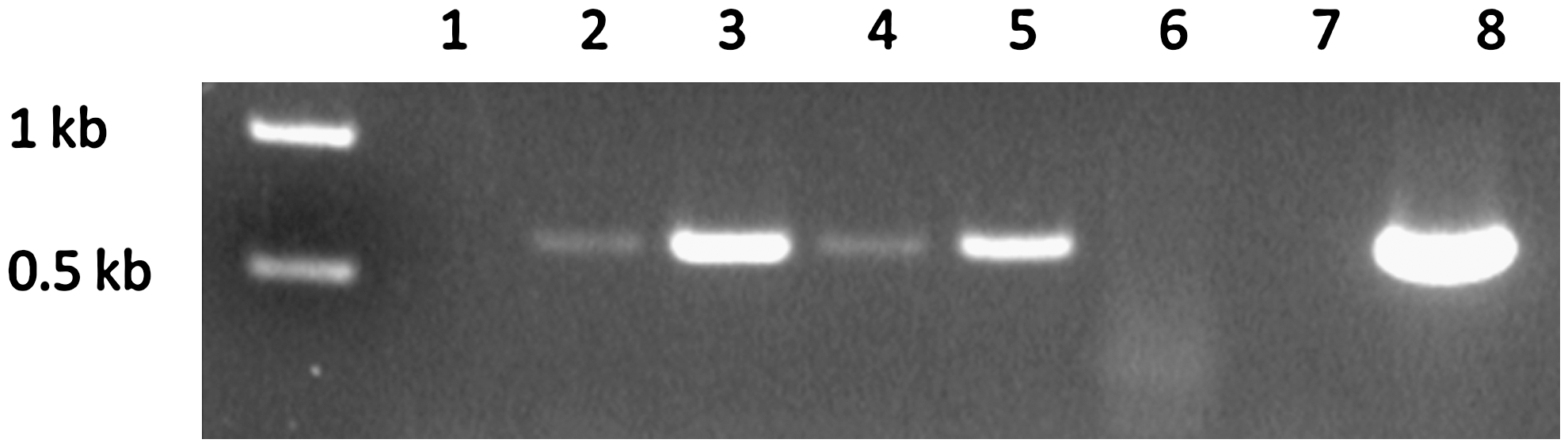
Polymerase chain reaction of the Borrelia burgdorferi 611 bp flaB amplicon from fecal DNA obtained during tick feeding time course. Lanes: (1) pre-tick attachment; (2) 30 h; (3) 56 h; (4) 76 h; (5) 96 h; (6) uninfected tick feces; (7) no DNA control; (8) B. burgdorferi B31 genomic DNA. Molecular weight markers are shown on the left.
Culture of tick feces
All BSK-II cultures (n = 5) of fecal sample swabs were negative after 4 weeks of incubation, although the same swab samples were PCR positive. This result indicated that viable organisms were not present in the feces at the time of collection. It was not determined whether organisms shed in the feces were nonviable before leaving the tick, or whether the spirochetes simply died by dessication when exposed to the external environment before culture inoculation.
Discussion
There are few accounts in the literature that have addressed shedding of B. burgdorferi from infected ticks through feces. In an early study by Benach et al. (1987) Borrelia were not observed by dark-field microscopic examination of feces, and subsequent culturing of fecal pellets in BSK media proved negative. Gern et al. (1996) reported observing Borrelia in the feces of infected ticks by microscopy, although it was not determined if the organisms were viable. In this study, we demonstrate B. burgdorferi DNA associated with infected-tick feces by PCR, but were unable to cultivate organisms from the samples.
Ticks defecate mainly and probably exclusively throughout host feeding. During the uptake of a bloodmeal by the tick, B. burgdorferi present within the midgut initiate replication in preparation for migration to the host. Spirochetal replication proceeds exponentially, producing numbers hundreds-fold more than in the prefed tick. Therefore, it may not be surprising that organisms from the midgut could be shed rectally throughout the feeding process. Our results indicated that B. burgdorferi were shed from the tick during the 4 day feeding period. However, we were unable to culture organisms from the fecal samples, suggesting that either the Borrelia are nonviable when shed in the feces, or more likely, unable to survive in the environment outside the tick for an extended period. In the time course experiment, fecal collections were made within 18–24 h; therefore, we were unable to determine the length of time viable spirochetes could have survived once shed.
We did not undertake a microscopic examination for Borrelia in fecal samples because we felt that culturing was a more sensitive method to assess the presence of viable spirochetes. Further, Borrelia have been described as “obligate” pathogens or host-propagated bacteria, meaning that they are not known to exist or survive naturally outside the environs of a mammalian or arthropod host such as in water or soil (Seshu and Skare 2001). We suspected that once outside the tick there would be few if any viable organisms, and therefore would not be observable or recognizable due to dessication.
There was a consideration that borrelial DNA could have originated not from fecal matter but instead from ticks that did not feed and perhaps died within the capsule, eventually liberating B. burgdorferi postmortem. However, we do not consider this possibility a likely source of DNA contamination, as we accounted for 100% of the capsule-contained ticks from three mice whereby the fecal samples were PCR positive (Table 2). Although 1–2 ticks were unaccounted for in the other capsule feeds (excluding the 2 mice that detached the capsule), we attribute that loss to normal tick escape and destruction by mouse grooming.
Although we collected more samples of feces from ticks infected with non-WT spirochetes (i.e., the bba64-complement), we saw no evidence that genetically modified Borrelia had a different phenotype regarding tick shedding than WT. In point of fact, the samples from WT-infected ticks demonstrated identical results confirming this observation.
The results of this study demonstrating shedding of B. burgdorferi in the excrement of ticks during feeding have important ramifications for researchers investigating mechanisms of vector transmission. The findings presented here should alert investigators performing PCR analysis of samples from tick feeds, to the potential presence of B. burgdorferi from fecal contamination. Failure to account for this variable could result in incorrect interpretation of experimental data and conclusions.
Footnotes
Acknowledgments
The authors thank Marc Dolan and Joe Piesman for their tick expertise, Jeannine Petersen for helpful insights and comments, Barbara J.B. Johnson for reviewing the article, and members of the DVBD Animal Resources, Andrea Peterson, Lisa Massoudi, Verna O'Brien, and John Liddell, for their assistance.
Disclosure Statement
No competing financial interests exist.