
Abstract
Bartonella species comprise a genus of gram-negative, fastidious, intracellular bacteria that have been implicated in association with an increasing spectrum of disease manifestations in dogs and human patients. In this study, chronic canine and human disease, for which causation was not diagnostically defined, were reported by the breeder of a kennel of Doberman pinschers. In addition to other diagnostic tests, serology, polymerase chain reaction, and enrichment blood culture were used to assess the prevalence of Bartonella sp. infection in the dogs and their owner. From five dogs, Bartonella vinsonii subsp. berkhoffii genotype I, multiple Bartonella henselae strains, and a species most similar to Candidatus B. volans, a rodent-associated Bartonella sp., were amplified and sequenced from biopsy tissues, cerebrospinal fluid, or blood enrichment cultures. The owner was bacteremic with B. vinsonii subsp. berkhoffii genotype I, the same subsp. and genotype detected in one of her dogs. These results further emphasize the ecological complexity of Bartonella sp. transmission in nature.
Introduction
In conjunction with the recent identification of numerous, previously unknown Bartonella species, there has been a concurrent recognition of novel reservoir hosts and the identification of an expanding spectrum of transmission-competent arthropod vectors (Chang et al. 2000, 2002, Dehio et al. 2004, Chomel et al. 2009a, Minnick and Battisti, 2009). Although the extent to which persistent infection with various Bartonella sp. can occur in pet dogs is poorly understood, natural infection with Bartonella vinsonii subsp. berkhoffii can span months in duration (Kordick and Breitschwerdt 1998). After experimental infection, B. vinsonii subsp. berkhoffii was associated with suppression of humoral and cell-mediated immunity (Pappalardo et al. 2000a, 2001).
Sequencing of the 16S-23S intergenic spacer (ITS) region has identified four B. vinsonii subsp. berkhoffii genotypes (Kordick and Breitschwerdt 1998, Chang et al. 2000, Maggi et al. 2006, Breitschwerdt et al. 2007, Duncan et al. 2007a). A specific vector that transmits genotypes of this subspecies among wild and domestic canines has not been determined, though ticks are currently considered a likely vectorial candidate (Breitschwerdt and Kordick 2000, Billeter et al. 2008, Angelakis et al. 2010, Telford and Wormser 2010). Risk factors associated with the transmission of most Bartonella sp. to animals or human patients remain poorly understood. Although flea and tick exposure and residing in rural environments are defined risk factors associated with Bartonella henselae and B. vinsonii subsp. berkhoffii exposure in dogs (Pappalardo et al. 1997, Henn et al. 2005), other studies have suggested that animal contact may represent an important risk factor for exposure and/or infection with these Bartonella sp. in immunocompetent patients (Kumasaka et al. 2001, Breitschwerdt et al. 2007). Several studies have implicated direct transmission of Bartonella sp. from dogs to humans (Keret et al. 1998, Tsukahara et al. 1998, Kerkhoff et al. 1999, Chen et al. 2007). Although bacterial viability in saliva has not been confirmed, Bartonella DNA has been sequenced from dog saliva (Duncan et al. 2007a, Kim et al. 2009). Due to frequent, close contact between companion animals and their owners, pets may serve as a source of human infection, as natural reservoirs for arthropod transmission or as environmental sentinels for the detection of these bacteria (Kordick and Breitschwerdt 1998, Breitschwerdt and Kordick 2000).
The overall objective of this study was to determine whether infection with a vector-borne organism was contributing to chronic illness in a kennel of dogs or their owner. When testing for Anaplasma, Babesia, Ehrlichia, and Rickettsia sp. proved negative or inconclusive, the molecular prevalence of Bartonella sp. was determined. Five dogs were infected with B. vinsonii subsp. berkhoffii genotype I, multiple B. henselae strains, and a species most similar to Candidatus B. volans. B. vinsonii subsp. berkhoffii genotype I was isolated by blood culture from the owner using Bartonella alpha-Proteobacteria growth medium (BAPGM).
Materials and Methods
Dogs and kennel owner
A veterinarian, who moved from New York to Virginia in 1977 at 20 years of age, resided in Floyd, Roanoke, and Montgomery counties from 1981 to the present time. This individual maintained a kennel of Doberman pinschers as household pets. Most of the dogs were also used in a breeding program, from which puppies were sold or maintained as personal pets. Historically, all Doberman pinschers bred by this veterinarian between 1978 and 1995 were considered healthy, with no illnesses such as polyarthritis, lymphadenopathy, dermal lesions, or neurological manifestations occurring during this time frame. Shortly after relocation of her household and kennel in 1995, the dogs experienced a heavy tick infestation. Ectoparasite control measures were instituted and neither tick or flea infestations were subsequently considered problematic; however, during the ensuing years the veterinarian reported a spectrum of chronic, progressive, and unexplained medical problems in all of the adult dogs that she maintained as personal pets or breeding animals. In her professional opinion, the number, type, and severity of these problems were unprecedented and were in contrast to her experiences as a breeder. Because transmission of a vector-borne pathogen was suspected, because prior diagnostic testing was negative or inconclusive, and because antibiotic and other treatments were ineffective, the kennel owner requested testing for vector-borne pathogens by the North Carolina State University College of Veterinary Medicine Vector Borne Diseases Diagnostic Laboratory (NCSU-CVM-VBDDL). In collaboration with the owner and faculty at the Virginia-Maryland Regional College of Veterinary Medicine (VMR-CVM), serological and molecular testing for other vector-borne pathogens was performed between 1997 and 2010.
Historically, all dogs lived predominantly indoors and slept in the house with the owner. When the dogs were outdoors, they were confined to a large fenced enclosure that included an open field with access to surrounding wooded areas. Dogs had direct or indirect contact with numerous wild animals that frequented the enclosure, including rabbits, deer, moles, voles, wild turkeys, and gray squirrels. Dogs were also observed ingesting wild animal feces. In April 2006, the kennel owner became acutely ill and subsequently has experienced chronic disease manifestations up to the present time.
Serology-based assays
Serological testing was performed using immunofluorescent assays (IFA) for detection of antibodies against Rickettsia rickettsii, Ehrlichia canis, and Babesia canis as previously described (Kordick et al. 1999). The starting dilution was 1:16, with endpoint titers defined as the last dilution at which brightly stained organisms could be detected by fluorescence. An ELISA-based test kit (SNAP 3Dx or SNAP 4Dx; IDEXX Laboratories, Inc., Westbrook, ME) was also used to test for Anaplasma sp., Borrelia burgdorferi, and Ehrlichia sp. antibodies. B. vinsonii subsp. berkhoffii and B. henselae antibodies were determined following traditional IFA practices with fluorescein-conjugated goat anti-dog or human IgG. B. vinsonii subsp. berkhoffii genotype I (isolate 93-CO-1 from the NCSU-IPRL, ATCC #51672) and B. henselae (strain Houston 1, ATCC #49882) were passed from agar-grown cultures of each organism into DH82 (a continuous canine histiocytic cell line) cultures. Heavily infected cell cultures were spotted onto 30-well Teflon-coated slides, air-dried, acetone-fixed, and stored frozen. Serum samples were diluted in phosphate-buffered saline containing normal goat serum, Tween-20, and powdered nonfat dry milk to block nonspecific antigen binding sites. Patient sera were screened at dilutions of 1:16 to 1:64. All sera that remained reactive at a titer of 1:64 were further tested with twofold dilutions out to a final dilution of 1:8192. A standard cut-off titer of 1:64 was defined as being seropositive.
BAPGM diagnostic platform
The BAPGM diagnostic platform incorporates DNA amplification of Bartonella sp. in three independent steps, as described previously (Maggi et al. 2005, Duncan et al. 2007b). Two milliliters of aseptically obtained blood, serum, or cerebrospinal fluid (CSF) samples was inoculated into 10 mL of BAPGM and cultured for 7 days (Duncan et al. 2007b). An un-inoculated BAPGM culture flask was processed simultaneously and in an identical manner with each batch of patient samples tested.
Tissue processing and DNA extraction
For fresh, frozen, or paraffin-embedded tissues, and blood samples, DNA was extracted for polymerase chain reaction (PCR) testing using the QIAmp mini kit and protocol provided by Qiagen (Valencia, CA). Cross contamination during necropsy collection or processing of paraffin-embedded tissues has previously been described (Varanat et al. 2009a); therefore, embedded samples were cut from the blocks using a new sterile scalpel blade, and for each processed block, a new paper work surface was used to obtain tissues from the block. All DNA samples were eluted in nuclease-free water and stored at −20°C until use.
Bartonella genus ITS region PCR
PCR screening for Bartonella DNA from blood, serum, fluids, tissues, and BAPGM cultures was performed using two primer sets in two individual PCRs targeting the ITS region located between the 16S and 23S rRNA genes, using oligonucleotides 325s-1100as as described previously (Duncan et al. 2007b) and 438s-1000as (438s: 5′-GGT TTT CCG GTT TAT CCC GGA GGG C-3′ and 1000as: 5′-CTG AGC TAC GGC CCC TAA ATC AGG-3′). The annealing temperature for primer amplification was 68.5°C with 55 cycles. The detection limit for these PCR assays is 2.5 genomic copies per reaction. A B. henselae Houston 1 strain type was used as a positive control throughout the study. All PCR and BAPGM culture-negative controls remained negative throughout the study.
Sequencing of ITS region amplicons
All PCR products were sequenced directly or after cloning as previously described (Diniz et al. 2007). Bacterial species and strain were defined by comparing similarities with other sequenced bacteria deposited in the GenBank database using the Basic Local Alignment Search Tool (Blast version 2.0).
Results
The veterinarian first sought consultation with the NCSU-CVM-VBDDL in 1998 because Dogs 1 and 2 had developed illnesses of unknown cause. Dogs 3–5 became ill after the breeder relocated to a new home within the same county in 2001. Dogs 1–2 died during the course of this investigation, whereas dogs 3–5 continue to live with the owner. The case designation, year of birth, sex, date of onset of illness, sample date providing the DNA evidence of Bartonella infection, predominant clinical abnormalities, and diagnoses for the dogs and owner are summarized in Table 1. Because testing by commercial laboratories had identified antibodies to R. rickettsii antigens, our initial diagnostic efforts focused on spotted fever group (SFG) Rickettsia serology and PCR testing. When these efforts generated no consistent indication of exposure to or infection with rickettsiae, diagnostic efforts were redirected to assess the prevalence of other vector-borne pathogens and Bartonella sp. Patient samples were tested retrospectively and prospectively.
Clinical abnormalities: F, fever; P, polyarthritis; S, swollen joints/extremities; I, inflammatory bowel disease; L, lymphadenopathy; D, dermal lesions; E, epididymitis; G, glomerulonephritis; N, neurological manifestations; H, hamartoma; PN, panniculitis; C, chest pain.
Bvb, Bartonella vinsonii subsp. berkhoffii; PCR, polymerase chain reaction.
Based upon PCR amplification and DNA sequencing, active infection with a Bartonella sp. was confirmed in blood, cerebrospinal fluid, or tissue biopsy samples derived from the owner and from Dogs 1 to 5. At no time point were any of the five dogs seroreactive to B. henselae or B. vinsonii subsp. berkhoffii antigens, whereas the owner was minimally seroreactive to B. henselae and B. vinsonii subsp. berkhoffii antigens.
Dog 1
In 1997, upper small intestinal biopsies were obtained endoscopically at VMR-CVM from Dog 1 due to suspected inflammatory bowel disease. Idiopathic polyarthritis was subsequently diagnosed in 1998, and in 2000, an acanthomatous epulis was biopsied. In 2007, the dog developed hypothyroidism and aspiration pneumonia secondary to megaesophagus, which persisted until 2009 when the dog was euthanized due to recurrent bouts of aspiration pneumonia. Retrospective Bartonella PCR testing in 2010 was performed on DNA extracted from formalin-fixed, paraffin-embedded tissue samples of the intestinal biopsy taken in 1997. The ITS sequence obtained from this amplicon was 99.5% (210/211 bp) similar to B. henselae (DQ529247); however, the strain could not be confirmed due to the short length of the clean, readable sequence. Bartonella DNA was not amplified from the acanthomatous epulis sample obtained in 2000.
Dog 2
Dog 2 was healthy until 1998, when cutaneous draining lesions and thrombocytopenia were documented. In September, the dog was reportedly seroreactive to rickettsial antigens with an endpoint titer of 1:64; however, repeat testing in October 1998 at the NCSU-VBDDL revealed no R. rickettsii seroreactivity. Retrospective PCR testing for SFG Rickettsia and Bartonella DNA using stored blood from October 1998 was also negative as well as blood and lymph node samples collected in July 2000. In 2007, B. henselae DNA was amplified and sequenced from surgically obtained frozen liver tissue removed at VMR-CVM in July of that year to investigate biochemical and sonographic evidence of a hepatopathy. The ITS amplicon was 99.4% similar (626/630 bp) to B. henselae strain SA2 (AF369529), excluding a 15 bp insertion at the 3′ end. It was also 99.5% homologous (602/605 bp) with a cat isolate obtained in our laboratory in 1995 (Isolate 95FO93), which contained the same 15 bp insertion at the 3′ end. At the time of the liver biopsy, a cutaneous histiocytic sarcoma was also surgically removed. By retrospective Bartonella ITS PCR in 2010, two bands were amplified from the paraffin-embedded histiocytic sarcoma (Fig. 1). Each band was excised and successfully sequenced following gel extraction. One band was 99.8% homologous (487/488 bp) with B. henselae Cal-1 (AF369527), whereas the other band was 99.2% homologous (514/518 bp) with B. henselae strain SA2 (AF369529). Despite treatment with azithromycin and morbofloxacin for 4 weeks, Dog 2 died in 2007 due to complications associated with glomerulonephritis.
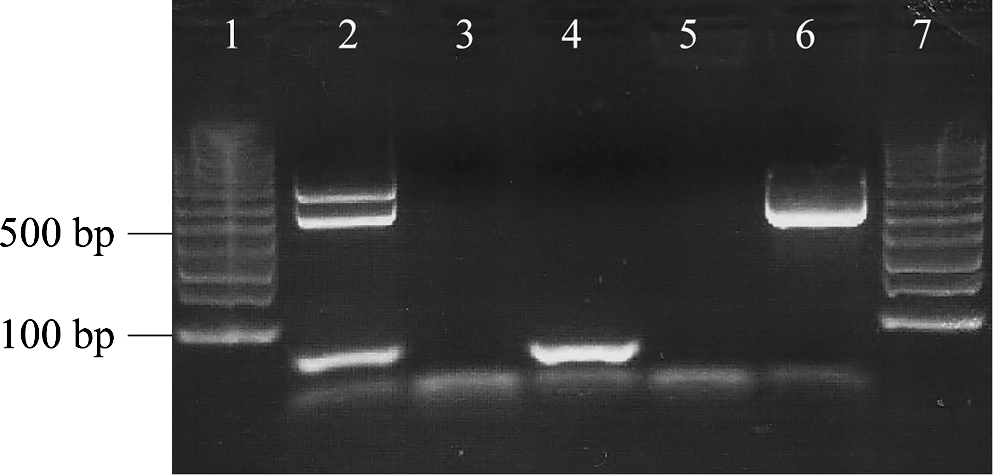
Agarose gel electrophoresis illustrating Bartonella genus intergenic spacer (ITS) polymerase chain reaction (PCR) amplicons (lane 2) obtained from the paraffin-embedded histocytic sarcoma from Dog 2. By sequencing, the top band contained Bartonella henselae Cal-1 DNA and the bottom band contained B. henselae SA2 DNA. Lanes 3 and 4: PCR-negative tissue extractions. Lane 5: PCR-negative control (specific pathogen-free dog DNA). Lane 6: PCR-positive control (B. henselae Houston 1 strain). Lanes 1 and 7: 1-Kb molecular marker.
Dog 3
Bartonella PCR was performed retrospectively using a surgically resected paraffin-embedded fibrosarcoma removed from the ventral thorax in 2005. An amplicon was 99.8% (446/447 bp) similar to B. henselae strain Houston 1 (NC005956). In early 2005, following a trip to Texas, both Dog 3 and Dog 4 developed dermal lesions on their muzzles, presumed to be histiocytomas. Shortly after this time, both dogs developed lesions on hocks, which were diagnosed as hamartomas. Samples from dermal lesions from Dog 3 were not available for Bartonella PCR. In January 2007, Dog 3 was referred to NCSU-VTH for shifting leg lameness, painful joints, and lymphadenopathy. Testing at this time revealed no seroreactivity to R. rickettsii, B. henselae, or B. vinsonii subsp. berkhoffi in serum, and blood was PCR negative for SFG Rickettsia and Bartonella DNA. BAPGM enrichment blood culture was repeated in December 2007, and a Bartonella sequence, not previously obtained in our laboratory or found in the GenBank database, was amplified. This ITS sequence had an 18 bp deletion at the 3′ end. After excluding this deletion, the remaining sequence was most similar (93% homology, 432/465 bp) to Candidatus Bartonella volans (EU294521), previously isolated from Southern flying squirrels. The sequence was 99% similar (482/487 bp) to a sequence obtained in our laboratory from a sea otter in 2008 (Carrasco et al. 2008). In February 2008, the dog was referred to the NCSU-VTH Neurology Service due to neck pain, ataxia, and hindlimb paresis. By magnetic resonance imaging there was evidence of cervical spondylomyelopathy at C5-C6 and the caudal cervical paraspinal muscles were hyperintense, suggestive of myositis or denervation atrophy. Cytological and protein analyses of cisternal cerebrospinal fluid were normal; however, B. vinsonii subsp. berkhoffii genotype I was PCR-amplified and sequenced from the CSF fluid sample. The CSF sequence was 99.8% similar (587/588 bp) to B. vinsonii subsp. berkhoffii genotype I (AF167988). The dog was not seroreactive to R. rickettsii, B. henselae or B. vinsonii subsp. berkhoffii antigens. PCR for SFG Rickettsia DNA was negative in blood and CSF samples. After treatment with azithromycin and rifampin for 6 weeks, neurological signs resolved and the dog has remained healthy.
Dog 4
In early 2005, Dog 4 developed dermal lesions on her muzzle, presumably histiocytomas, similar to those seen on Dog 3. Later in 2005, following surgical removal of a cutaneous mass, Dog 4 was found to have a cutaneous follicular hamartoma. DNA was extracted from a stored paraffin-embedded hamartoma tissue sample, and ITS PCR generated an amplicon that was 99.8% (487/488 bp) similar to B. henselae SA2 (AF369529). In August 2006, the dog developed tremors and twitching of the distal extremities of undetermined cause. Between September and December 2006, splenomegaly accompanied by mild to moderate thromobocytopenia and mild anemia were repeatedly documented. When incessant chewing at the elbows and aberrant behavior (inattention and staring at the elbows) developed, Dog 4 was treated with doxycycline for 6 weeks. In December 2006, PCR from blood was negative for SFG Rickettsia and Bartonella sp. Serum was not provided. Serum obtained in December 2007 was reactive by IFA testing to E. canis antigens (titer 1:256) but was negative for Ehrlichia by SNAP 4DX testing and weakly SNAP positive to Anaplasma antigens (A. phagocytophilum or A. platys). Repeat testing in January 2008 yielded only seroreactivity to Anaplasma by SNAP 4DX testing, most likely reflecting seroconversion to A. phagocytophilum. There was no serological or PCR evidence of infection with Bartonella or SFG Rickettsia between 2006 and 2009.
Dog 5
In 2005, Dog 5 was found to have an idiopathic peripheral neuropathy at VMR-CVM, in conjunction with coincident seroreactivity to B. burgdorferi. In August 2006, a large, firm, moveable dorsal thoracic mass containing 12 mL of serosanguineous fluid was aspirated. Cytological findings included a large number of macrophages (68%) with accompanying erythrophagocytosis, nontoxic neutrophils (17%), lymphocytes (14%), rare eosinophils (1%), and no visualized organisms. A complete blood count was normal and aerobic, anerobic, and fungal cultures of the aspirated fluid were negative. Due to continued drainage, the mass was surgically excised in October and the histopathological diagnosis was pyogranulomatous panniculitis. Repeat aerobic, anerobic, fungal cultures, and atypical mycobacterial cultures were negative. The dog was not seroreactive to B. henselae, B. vinsonii subsp. berkhoffii, or R. rickettsii antigens. After administration of doxycycline for 6 weeks, there was no additional palpable pathology in the dorsal thoracic region. Retrospective testing of surgically obtained paraffin-embedded tissue from the mass was PCR positive for B. henselae (Fig. 2). The ITS sequence was 99.6% (516/518 bp) similar to B. henselae SA2 (AF369529).
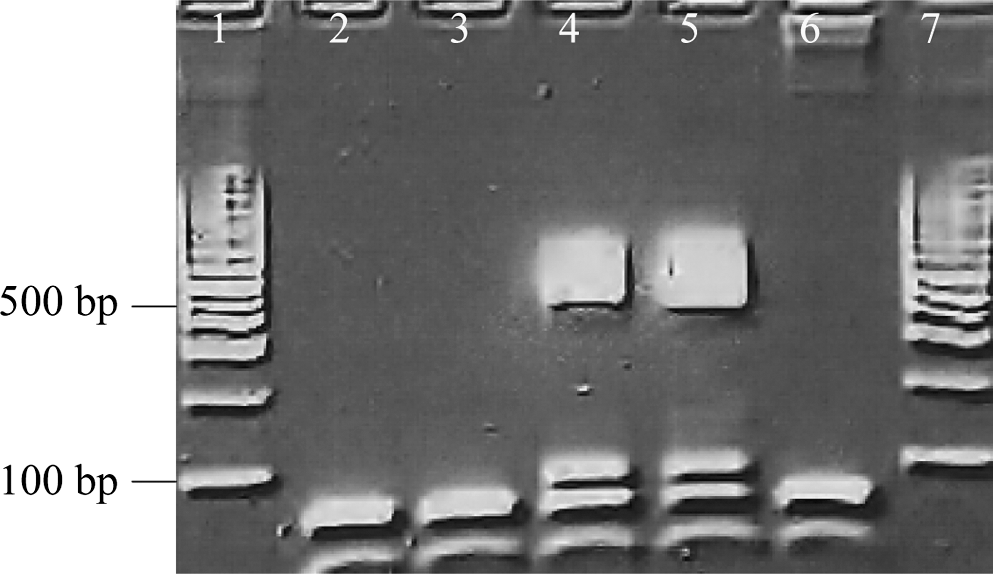
Agarose gel electrophoresis illustrating a Bartonella genus ITS PCR amplicon (lane 4) obtained from the paraffin-embedded panniculitis lesion from Dog 5. DNA sequencing of the amplicon confirmed B. henselae (SA2 strain). Lanes 2 and 3: PCR-negative tissue extractions. Lane 5: PCR-positive control (B. henselae Houston 1 strain). Lane 6: PCR-negative control (specific pathogen-free dog DNA). Lanes 1 and 7: 1-Kb molecular marker.
Owner
Beginning in April 2006, the female veterinarian, who bred and owned the Doberman pinschers described in this investigation (Dogs 1–5), developed progressive neurological disabilities, experienced chronic joint and muscle pain, and was found to have a sensory-motor neuropathy. Five months after the onset of illness, she was barely able to walk or use her hands and arms. In addition to occupational exposure in her practice, this veterinarian reported frequent close contact with her dogs, including unprotected exposure to their saliva and infrequent minor scratch or bite wounds. In September 2006, after a 5-month history of progressive illness, the veterinarian was found to have Lyme disease, after which she was treated with doxycycline for 4 weeks. As there was no resolution of symptoms after doxycycline treatment, she was subsequently treated with 4 weeks of intravenous ceftriaxone, which resulted in overall symptomatic improvement; however, she was classified as neurologically disabled and continued to experience symptoms, including continuous pain, swelling and edema in her extremities, chest pain, and dermal lumps and plaques. In December 2007, the veterinarian was entered into an Institutional Review Board (IRB)-approved research study (NCSU IRB #164-08-05) designed to assess the occupational risk of Bartonella sp. infection in animal health professionals. Using Bartonella ITS primers, B. vinsonii subsp. berkhoffii genotype I was amplified from a BAPGM enrichment blood culture. The amplicon was 100% homologous (659/659 bp) with B. vinsonii subsp. berkhoffii genotype I (AF167988). By IFA testing, seroreactivity to B. vinsonii subsp. berkhoffii and B. henselae antigens was not detected. B. vinsonii subsp. berkhoffii genotype I DNA was again amplified following direct extraction of a serum sample obtained in January 2008. ITS PCR from the blood sample was also positive, but the amplicon was not successfully sequenced. After enrichment in BAPGM for 14 days, B. vinsonii subsp. berkhoffii genotype I was sequenced from an agar plate subculture isolate. Sequences of PCR products obtained from the serum and the subculture isolate, which was obtained weeks later, were 100% homologous with B. vinsonii subsp. berkhoffii genotype I (AF167988). At this time, the patient was seroreactive to B. henselae and B. vinsonii subsp. berkhoffii genotypes I, II, and III at titers of 1:32, 1:16, 1:64, and 1:64, respectively. There was no postantibiotic PCR evidence of Bartonella infection in February 2008, when blood and serum were again tested using the BAPGM platform; however, similar antibody titers to those obtained from samples in January 2008 were obtained to the respective Bartonella antigens.
Discussion
In this study, we describe molecular evidence of Bartonella sp. infection in an owner and five adult dogs maintained in her household. From an ecological perspective, numerous domestic and wild animals have coevolved with various Bartonella sp. and with the vectors that transmit these bacteria among animal populations. Vector preference generally confines transmission of a given Bartonella sp. among defined animal populations, such as cats, cows, coyotes, foxes, or rodents; however, inadvertent transmission to incidental hosts, such as pet dogs or humans, can occur with resulting pathology.
All five Bartonella PCR-positive dogs were infected with various ITS strains of B. henselae, which most likely reflects exposure to multiple strains of this species or, alternatively, changes in the 16S-23S ITS region may have occurred during in vivo infection of the host (Diniz et al. 2007, Oliveira et al. 2010). Precautions were taken to prevent DNA contamination or carryover during sample processing. Contamination of tissue samples with Bartonella DNA can occur during necropsy or histological tissue processing (Varanat et al. 2009a); therefore, in this study, sterile scalpel blades were used to manually cut patient tissues from individual paraffin blocks. All BAPGM enrichment cultures and PCR procedures were conducted under stringent conditions to avoid PCR amplicon contamination, and all negative controls remained negative throughout the course of this study.
Due to current limitations in knowledge regarding Bartonella sp. vector transmission, the long time span of this study, and the intermittent and uncontrolled timing of diagnostic testing of these dogs and their owner, it is not possible to determine the source of Bartonella infections or mode(s) of transmission. For Dog 2, three distinct ITS strains of B. henselae were sequenced from liver or the histiocytic tumor tissue. One strain was most similar to B. henselae Cal-1, the second strain to B. henselae SA2, and the third was an SA2-like strain. In contrast, Dog 3 was infected with B. henselae, B. vinsonii subsp. berkhoffii genotype I, and a novel B. volans–like strain at different time points between 2005 and 2008. For the owner and Dog 3, enrichment culture in BAPGM was required before PCR was able to confirm B. vinsonii subsp. berkhoffii genotype I infection in the owner and B. henselae strain Houston 1 infection in the dog, respectively. Previous studies have supported enhanced diagnostic sensitivity associated with this liquid enrichment culture approach in dogs and human patients (Maggi et al. 2005, Duncan et al. 2007b, Kosoy et al. 2008, 2010, Breitschwerdt et al. 2010a, 2010b).
Both the veterinarian and Dog 3 may have been infected by a common environmental source of B. vinsonii subsp. berkhoffii, as the same ITS genotype sequence was amplified from the dog's cerebrospinal fluid and the veterinarian's BAPGM blood culture and subculture isolate. Several recent studies support an increased likelihood of Bartonella exposure and/or infection in immunocompetent people with extensive animal and arthropod contact (Kumasaka et al. 2001, Breitschwerdt et al. 2007, 2008). B. vinsonii subsp. berkhoffii genotype I bacteremia and seroconversion were recently reported in a veterinarian after experiencing a needle stick, which occurred while aspirating a cutaneous histiocytic tumor in a dog who subsequently was found to be infected with B. vinsonii subsp. berkhoffii (Oliveira et al. 2010). In addition, Bartonella sp. DNA has been detected in dog saliva (Duncan et al. 2007a, Kim et al. 2009), and direct bite transmission of Bartonella sp. from dogs to humans has been suggested (Keret et al. 1998, Tsukahara et al. 1998, Kerkhoff et al. 1999, Chen et al. 2007). In light of evolving information, the possibility of zoonotic transmission of Bartonella sp. from dog to owner in this study cannot be discounted due to their close interactions. As of yet, no definitive mode of transmission for any Bartonella sp. to a dog has been established (Billeter et al. 2008). Regardless of the mode(s) of transmission, B. vinsonii subsp. berkhoffii has become an important, emerging zoonotic pathogen. Disease development may be more likely when dogs become immunocompromised (Yager et al. 2010) or when opportunistic infection becomes established in a nonreservoir host such as a cat, dog, or human (Breitschwerdt et al. 2009b, 2010, Breitschwerdt and Maggi, 2009, Varanat et al. 2009b).
Based upon the large number of rodent-adapted Bartonella sp. found in various urban and rural ecosystems throughout the world, including several rodent species that are frequently sold as pets (Inoue et al. 2009), it is likely that rodent-reservoir Bartonella sp. infection of dogs and people is a more frequent occurrence than is currently appreciated (Kosoy et al. 2008, 2010). A high prevalence of Bartonella bacteremia (42.2%) has been reported in rodent surveys from the southeastern United States (Kosoy et al. 1997); therefore, dogs in frequent contact with rodents, as was the case in this study, may become infected with a variety of rodent Bartonella species, for which the pathogenic potential is currently unknown. Previously, Bartonella washoensis and Bartonella elizabethae, with the California ground squirrel (Spermophilus beecheyi) and the rat (Rattus sp.) as the associated reservoirs, respectively, have been shown to infect dogs and people (Mexas et al. 2002, Chomel et al. 2003, Marié et al. 2006). In addition, we have described infection with a novel Bartonella strain, most closely related to Candidatus Bartonella volans, in an 86-year-old farmer who experienced several postoperative complications, including Candida albicans esophagitis, hallucinations, seizures, and encephalopathy after surgery for a fractured femur (Breitschwerdt et al. 2009a). That strain was similar (98.9%, 460/465 bp) but not identical to the BAPGM enrichment blood culture Bartonella sequence detected in Dog 3. Currently, the reservoir(s), mode of transmission, and putative arthropod vector(s) for these novel B. volans–like strains are unknown.
As noted in other studies, there were discrepant results between Bartonella PCR results and seroreactivity to the Bartonella test antigens in the current study. A study from our laboratory documented Bartonella infection in dogs that lacked a detectable humoral immune response by IFA testing (Duncan et al. 2007b). When testing human sera, varying antigenic expression between B. henselae strains also appears to contribute to false-negative B. henselae IFA results (Drancourt et al. 1996). In addition, several case reports have demonstrated detection and/or isolation of Bartonella in seronegative human patients (Raoult et al. 1994, Drancourt et al. 1996, Brouqui et al. 1999, Ehrenborg et al. 2000).
On an evolutionary basis, Bartonella sp. are highly adapted, vector-transmitted, erythrocytic, and endotheliotropic bacteria; therefore, satisfying Koch's postulates for disease causation has been challenging in both human and veterinary medicine (Jacomo et al. 2002, Chomel et al. 2009a). Prospective studies are needed to better define the duration of bacteremia and the humoral response after natural infection and to determine if Bartonella species play a role in dog and human patients with chronic disease manifestations, including polyarthritis, glomerulonephritis, and peripheral neuropathies. Intermittent sampling for over a decade did not allow us to establish the onset or modes of Bartonella sp. transmission, but DNA of an intriguing number of Bartonella sp. and strains were detected in tissues, CSF, and blood enrichment cultures over time. Our goal in reporting these results is not to infer disease causation for the owner or for her dogs but to further emphasize the ecological complexity of the genus Bartonella in nature.
Footnotes
Acknowledgments
We thank the veterinarian, who was also the kennel owner, for providing historical information relative to her dogs and herself and for her reading of the article before submission. We also thank members of the Vector Borne Disease Diagnostic Laboratory at North Carolina State University for diagnostic testing data generated on these dogs. This research was supported in part by a grant from the American College of Veterinary Internal Medicine Foundation and Bayer Animal Health. Natalie Cherry is the Novartis Fellow in Vector Borne Infectious Diseases.
Disclosure Statement
In conjunction with Dr. Sushama Sontakke and North Carolina State University, Dr. Breitschwerdt holds U.S. Patent No. 7,115,385; Media and Methods for cultivation of microorganisms, which was issued October 3, 2006. He is the chief scientific officer for Galaxy Diagnostics, a newly formed company that provides diagnostic testing for the detection of Bartonella species infection in animals and in human patient samples. Dr. Ricardo Maggi is the Scientific Technical Advisor and Laboratory Director for Galaxy Dx. The remaining authors have no potential conflicts of interest.