
Abstract
During summers in 2007 and 2008, an investigation was conducted to identify the distribution of mosquitoes and circulation of mosquito-borne arboviruses in Inner Mongolia, China. A total of 10,542 mosquitoes consisting of seven species from the Aedes, Culex, and Anopheles genera were trapped by UV-light traps, and they were sorted into 211 pools according to species, location, and date of collection. The result showed that Aedes dorsalis was the most common species, accounting for 41.0% (4327/10,542) of the total. Culex modestus (17.1%, 1801/10,542) made up the second largest species, followed by Aedes flavidorsalis (16.3%, 1714/10,542). Six virus isolates were obtained from pooled mosquitoes using cell culture and were identified as Tahyna virus (two isolates from Ae. dorsalis and C. modestus), Banna virus (one isolate from C. modestus), and Culex Pipiens pallens Densovirus (three isolates from Aedes caspius) by serological and molecular methods.
Introduction
Because of the close contacts between residents and livestock, zoonotic diseases frequently occur in local areas. Tick-borne encephalitis has been reported from Daxinganling forest of Inner Mongolia (Lu et al. 2008, Gao et al. 2010). Borrelia burgdorferi, spotted fever group Rickettsia, and Babesia microti were also detected from ticks collected in the ports of Inner Mongolia (Hao et al. 2009). From 1960s to 1980s, Japanese encephalitis prevailed in Inner Mongolia, where the incidence rate of Japanese encephalitis was up to 9.66/100,000 in 1974 (Wang et al. 2009, Cao et al. 2010).
With economic development over the past 30 years, Inner Mongolia has experienced extensive increases in urbanization, population growth, and commerce. Information regarding transmission of arboviruses in the region is limited. In this study, we collected mosquitoes in selected areas across Inner Mongolia during the summers of 2007 and 2008 to provide preliminary information regarding distribution of mosquitoes and mosquito-borne arboviruses in the region.
Materials and Methods
Mosquito collections
The mosquitoes were collected in 12 sites in four districts in Inner Mongolia. The collection sites were located in the east (six sites in Tongliao), middle (three sites in Huhhot), and west (two sites in Bayannaoer and one site in Wuhai) of Inner Mongolia (Fig. 1). Areas from the north frontier to Tongliao (900 km in distance) possess a great deal of forest (Daxinganling primeval forest) seldom with human activities. Areas from the west boundary to Wuhai and Bayannaoer (700 km in distance) are mainly Gobi desert with few people living there. Therefore, the four sample districts in our study, namely Tongliao in the east, Huhhot in the middle, and Bayannaoer and Wuhai in the west, almost covered main areas where most of people lived by agriculture and livestock husbandry. The sites were placed in agricultural villages containing 2000–4000 people and with pigsties, cattle farms, and ponds.
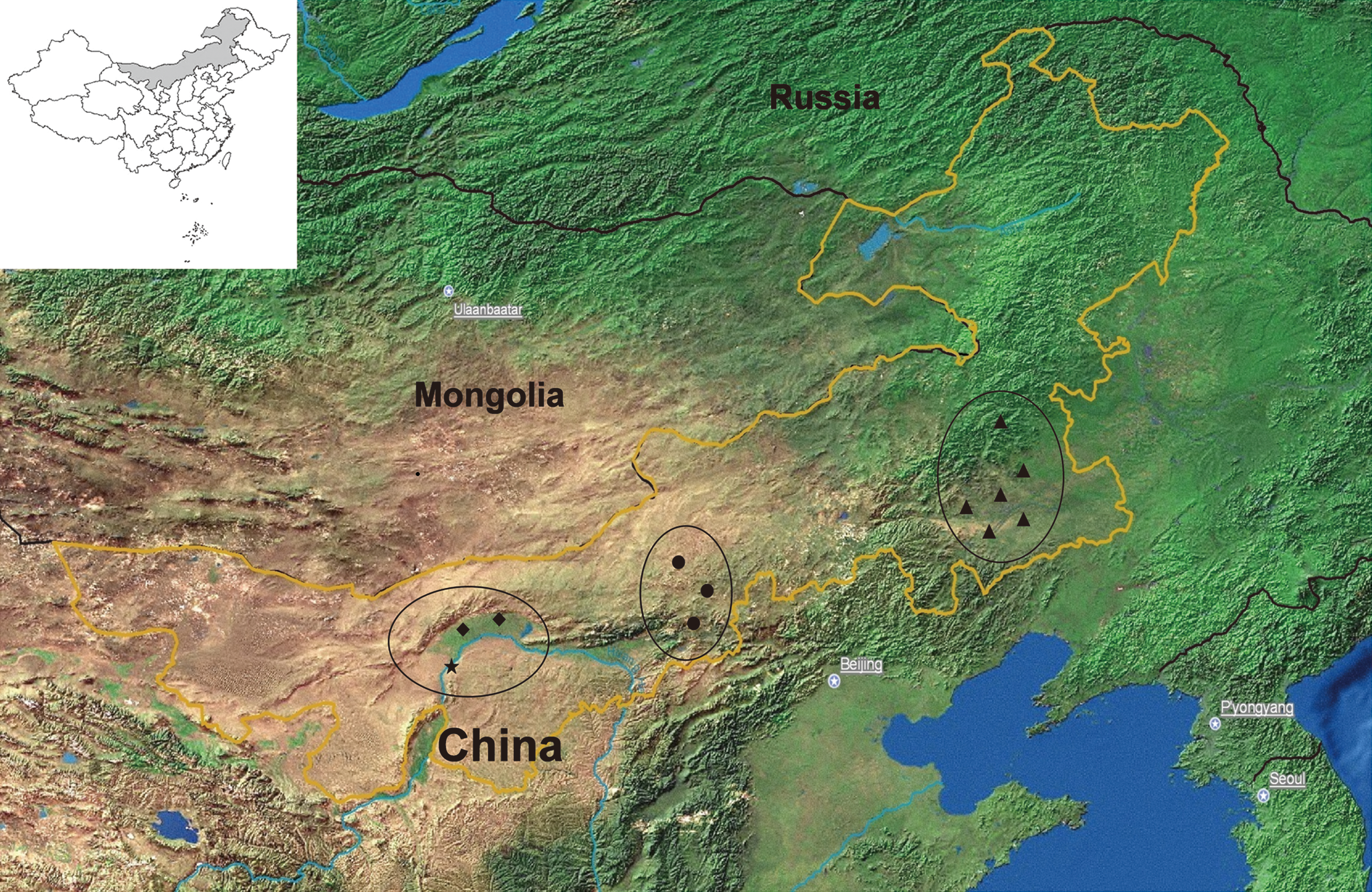
Map of Inner Mongolia showing the location of different sites studied (triangle: six sites in Tongliao; dot: three sites in Huhhot; diamond: two sites in Bayannaoer; star: one site in Wuhai). The yellow line represents the border of Inner Mongolia. Map of China (upper left) showing the location of Inner Mongolia in China.
During August 1st to 21st in 2007 and July 29th to August 26th in 2008, adult mosquitoes were collected using UV-light traps (Hubei Lucky Star Environment Protection Co., Ltd.). The traps were placed before sunset and mosquitoes were collected from the traps in the following morning after sunrise. Adult female mosquitoes were identified to species by morphologic keys (Lu 2003, Tian 2009) and pooled into groups by species, date, and site of collection. Approximately 50 mosquitoes were placed in each pool (Gu and Novak 2004). Pools were transported to the laboratory on dry ice and stored at −80°C until tested.
Virus isolation and identification
Mosquito pools were triturated in minimum essential medium supplemented with 2% fetal bovine serum, 1% glutamine, 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 1 μL/mL of fungizone using TissueLyser apparatus (Qiagen) at 4°C for 3 min. The suspensions were clarified by centrifugation at 4°C and 12,000 rpm for 10 min (Bryant et al. 2005). Clarified homogenates were inoculated into monolayers of BHK-21 and C6/36 cells and incubated at 37°C and 28°C, respectively. Cells were observed daily for signs of cytopathic effects (CPEs) from day 1 through 7 postinfection.
The indirect immunofluorescence assay was conducted with a panel of arboviruses antisera with fluorescein isothiocyanate-conjugated goat anti-mouse IgG as the second antibody (Sigma) by previously described procedures (Li et al. 2010). The panel of antisera included broadly reactive antisera against alphaviruses, broadly reactive antisera to bunyaviruses, specific antisera to Getah virus, antisera to Sindbis virus, antisera against Japanese encephalitis virus, and antisera against Banna virus prepared by our laboratory (Liang et al. 1993, Tao et al. 2002). Immune ascites against the prototype Tahyna virus (TAHV) (Bardos 92 strain) provided by Centers for Disease Control and Prevention (CDC) were also used.
Plaque reduction neutralization test (PRNT) was conducted as previously described (Li et al. 2010). The isolate with immune ascitic fluid (TAHV Bardos 92 stains; CDC) was tested in PRNT. The neutralizing antibody titer was identified as the highest ascitic fluid dilution that reduced the number of virus plaques by 90% in the test.
Molecular identification
Extraction of viral RNA was done with cell culture supernatants using the QIAamp Viral RNA Mini Kit (Qiagen), and first-strand complementary DNA was synthesized using the Ready-to-Go™ You Prime First Strand Beads (GE Healthcare) according to the manufacturer's protocol. Polymerase chain reaction (PCR) was conducted with primers designed for generic amplification of bunyaviruses (Kuno et al. 1996), flaviviruses (Kuno et al. 1998), and alphaviruses (Bryant et al. 2005) as well as species-specific primers for identification of TAHV (Lu et al. 2009) and Culex pipiens pallens Densovirus (CppDNV) (Zhai et al. 2008) (Table 1). Amplified DNA fragments were visualized on 1% agarose gels. Positive DNA fragments were extracted using the TaKaRa DNA fragment Purification Kit (TaKaRa). The PCR fragments were sequenced using BigDye Terminator V3.1 cycle sequencing kit (PE Applied Biosystems) and analyzed using ABI 3730 XL DNA Analyzer (PE Applied Biosystems).
CppDNV, Culex pipiens pallens Densovirus; TAHV, Tahyna virus.
Initial nucleic acid sequence assembly and analysis were conducted using the Seqman program (DNASTAR;
Results
Distribution of mosquitoes
A total of 10,542 mosquitoes consisting of seven species from three genera (Aedes, Culex, and Anopheles) were collected in 2 years, which included Aedes dorsalis, Aedes flavidorsalis, Aedes caspius, Aedes vexans, Culex modestus, C. pipiens pallens, and Anopheles sinensis (Table 2). Aedes mosquitoes accounted for 79.9% (8426/10,542) of all mosquitoes collected and Ae. dorsalis was the most common species accounting for 41.0% (4327/10,542) of the total, followed by Ae. flavidorsalis (16.3%, 1714/10,542), Ae. caspius (14.2%, 1493/10,542), and Ae. vexans (8.5%, 892/10,542). C. modestus accounted for 17.1% (1801/10,542) of the total. C. pipiens pallens (2.3%, 248/10,542) and An. sinensis (0.6%, 67/10,542) were rarely collected.
Ae., Aedes; Cx., Culex; An., Anopheles.
East includes six sites in Tongliao district; middle includes three sites in Huhhot district; west includes two sites in Bayannaoer and one site in Wuhai district.
%, the number of mosquitoes of collected species divided by the total number of mosquitoes in site.
Isolation of viruses
The 10,542 mosquitoes were sorted into 211 pools according to species, location, and date of collection. Pools contained ∼50 mosquitoes each, and all the pools were processed for virus isolation. A total of six isolates were obtained, which caused CPEs (Table 3). Isolates NM08003 and NM08010 caused CPE in 2 days only in BHK-21 cells but not in C6/36 cells. Isolates NM0706, NM0728, NM0740, and NM0741 caused CPE in C6/36 cells but not in BHK-21 cells. The onset of CPE by NM0706 isolate was 3 days after infection and CPE caused by NM0728, NM0740, and NM0741 isolates began 4 days after inoculation.
See the details about the identification of BAV in the paper by Cao et al. (2009).
BAV, Banna virus.
Identification of TAHV
Both NM08003 and NM08010 isolates reacted positively in indirect immunofluorescence assay tests with antisera against bunyaviruses and ascites against TAHV prototype (Bardos 92 strain, CDC), but not with antisera against alphaviruses, Sindbis virus, Getah virus, Japanese encephalitis, and Banna virus, indicating that both isolates were TAHV or a closely related bunyavirus. Subsequently, PRNT was performed on NM08003 isolate with ascites against TAHV. NM08003 isolate was identified to be TAHV, with a titer of 1:6400.
NM08003 and NM08010 isolates were RT-PCR positive when primers for bunyaviruses and TAHV were used and the S segment was sequenced. The result showed that the S segment of both isolates has 977 nt and the nucleotide sequence identity between them was 99.9%. The nucleotide sequence identity of NM08003 and NM08010 isolates with TAHV Bardos 92 strain (GenBank accession number: U47142), TAHV Xingjiang isolate (TAHV XJ0625, GenBank accession number: EU622820), and Qinghai isolates (QH07029 and QH07060 isolates) were 96.9%, 93.1%, 91.3%, and 91.3%, respectively.
The phylogenetic tree was constructed based on open reading frame sequence (coding for an N protein of 235 amino acid) of S segment. The cladogram showed that both NM08003 and NM08010 isolates were grouped into the clade of TAHV and were closely related to TAHV Bardos 92 strain, which suggested that these isolates are TAHV (Fig. 2).

Phylogenetic analysis of Tahyna virus isolates (NM08003 and NM08010) based on the open reading frame of the small segment. The tree was rooted by using Bunyamwera virus as the outgroup virus. The GenBank accession numbers of viruses are listed behind the viruses/strain.
Identification of CppDNV
PCR of NM0728, NM0740, and NM0741 isolates was performed with specific primers for CppDNV NS1 gene and positive fragments were sequenced. The alignment of the three isolates' nucleotides showed 99.9% homology. The sequences of three isolates matched CppDNV nucleic acid sequences in GenBank (CppDNV-XJ057, GenBank accession number: EF579760; CppDNV-YN0569, GenBank accession number: EF579765; CppDNV-XJ059, GenBank accession number: EF579759; CppDNV-JZ16, GenBank accession number: EF579756), with over 99% identity.
Discussion
Mosquito records for the past years in Inner Mongolia showed that mosquitoes belonging to 42 species from five genera were found, and they consisted of 28 species of Aedes (66.7%), 6 species of Culex (14.3%), 4 species of Culiseta (9.5%), 3 species of Anopheles (7.1%), and 1 species of Coquillettidia (2.4%) (Tian 2009). The mosquito species collected in this survey were included in the documents previously recorded. Further, our study indicated that Ae. dorsalis was the dominant mosquito species in the east and middle areas of Inner Mongolia, but Ae. caspius and Ae. flavidorsalis were the dominant mosquito species in Bayannaoer and Wuhai, respectively, west of Inner Mongolia in August for 2 consecutive years, 2007 and 2008.
In China, TAHV was first isolated from Culex mosquitoes collected in Kashi of Xinjiang Province, west of China, in 2006 (Lu et al. 2009) and later from Aedes detritus in Qinghai-Tibet Plateau in 2007 (Li et al. 2010). Moreover, TAHV antibody was detected in residents, cows, sheep, and swine in these locations, indicating that this virus circulated in these areas (Gu and Artsob 1987, Lu et al. 2009, Li et al. 2010). In this study, TAHV was isolated from mosquitoes in Inner Mongolia, suggesting a wide distribution of TAHV in the western and northwestern areas of China. TAHV, a member of California encephalitis sero group in the family Bunyaviridae, was first isolated from Ae. caspius collected in former Czechoslovakia in 1958 (Bardos and Danielova 1959). Since then, TAHV was isolated from several mosquito species, including Ae. caspius, Ae. vexans, Aedes cinereus, Aedes Ochlerotatus detritus, Aedes cantans, Aedes punctor, Aedes communis, Aedes flavescens, Aedes excrucians, Culiseta annulata, C. modestus, Culex pipiens, and Anopheles hyrcanus (Bardos and Danielova 1959, Hubálek 2008, Li et al. 2010). This is the first evidence of TAHV isolated from Ae. dorsalis.
Banna virus (Reoviridae, Seadornavirus) has been isolated from Culex tritaeniorhynchus, C. pipiens pallens, Culex annulus, Culex pseudovishnui, Ae. vexans, Ae. dorsalis, Aedes vagus, and An. sinensis in Yunnan, Gansu, Beijing, Shanxi, and Liaoning provinces in China (Tao and Chen 2005, Sun et al. 2009, Liu et al. 2010) as well as in Indonesia (Attoui et al. 2005) and Vietnam (Nabeshima et al. 2008). Our report of Banna virus isolated from C. modestus in Tongliao (42°85′N, 120°65′E) may represent the northmost area where Banna virus occurs. Banna virus has been isolated from a human in Xishuangbanna, Yunnan Province, China, suggesting that it infects man (Xu et al. 1990). However, this speculation requires further investigation.
In this study, three strains of CppDNV were obtained from Ae. caspius collected in Inner Mongolia. Their nucleotide sequence showed 99% similarity to CppDNV isolated from mosquitoes collected in Liaoning, Xingjiang, and Yunnan provinces and suggests that they are novel variants of viruses within the Brevidensovirus genus (Zhai et al. 2008).
This 2-year study showed that Aedes mosquitoes were the dominant mosquitoes in Inner Mongolia and that TAHV was isolated from Ae. caspius and C. modestus, which represents expansion of the geographic distribution of TAHV in China, after isolation of TAHV in Xinjiang Province (Lu et al. 2009) and Qinghai-Tibet Plateau (Li et al. 2010). Further investigation is needed to determine the disease burden caused by TAHV in this region.
Footnotes
Acknowledgments
The authors thank Dr. Roger S. Nasci (CDC) for his careful review of the article and valuable suggestions. The authors also thank staff members of CDC in Tongliao, Huhhot, Bayannaoer, and Wuhai, China, for the assistance in collection of mosquitoes. This work was supported by grants from the Ministry of Science and Technology, People's Republic of China (No. 2003BA712A08-01 and 2008ZX10004-001); Development Grant of State Key Laboratory for Infectious Disease Prevention and Control (2008SKLID105); and China–US CDC Cooperative Agreement U19-GH000004.
Disclosure Statement
No competing financial interests exist.