
Abstract
Seabirds act as natural reservoirs to Lyme borreliosis spirochetes and may play a significant role in the global circulation of these pathogens. While Borrelia burgdorferi sensu lato (Bbsl) has been shown to occur in ticks collected from certain locations in the North Pacific, little is known about interspecific differences in exposure within the seabird communities of this region. We examined the prevalence of anti-Bbsl antibodies in 805 individuals of nine seabird species breeding across the North Pacific. Seroprevalence varied strongly among species and locations. Murres (Uria spp.) showed the highest antibody prevalence and may play a major role in facilitating Bbsl circulation at a worldwide scale. Other species showed little or no signs of exposure, despite being present in multispecific colonies with seropositive birds. Complex dynamics may be operating in this wide scale, natural host–parasite system, possibly mediated by the host immune system and host specialization of the tick vector.
Introduction
Antibody (Ab) prevalence reflects both past and present exposure of hosts to parasites. A few studies have investigated the prevalence of anti-Borrelia antibodies in seabirds (Gauthier-Clerc et al. 1999, Gylfe et al. 1999, Gasparini et al. 2001, Staszewski et al. 2008), but the number of species and locations considered in these studies is often limited. In the North Atlantic, significant spatial and interspecific variability in exposure to spirochetes has been reported (Staszewski et al. 2008), and the anti-Borrelia Ab levels of individual birds was shown to be highly stable between years in one wide-spread species, the black-legged kittiwake Rissa tridactyla (Staszewski et al. 2007). In the North Pacific, the presence of Borrelia in ticks has been reported (Olsen et al. 1995, Comstedt et al. 2009, Gómez-Díaz et al., 2011), but little is known about variability in Bbsl exposure within and among seabird communities. North Pacific seabird communities are composed of a rich array of colonial species, and share with the North Atlantic communities both the tick vector I. uriae and some seabird host species (black-legged kittiwake, common murre Uria aalge, and thick-billed murre Uria lomvia). Among the diversity of seabirds that make up these communities, there are several species of particular interest in terms of tick and pathogen exposure; two sister species pairs of seabirds, namely, red-legged Rissa brevirostris and black-legged kittiwakes, and common and thick-billed murres, that breed in similar habitats and may share local tick populations and have similar exposure levels. Likewise, there are also a series of Aethia auklet species, some of which have been suggested to use an odorant as a defence against ticks (Douglas et al. 2004), but whose infestation levels are largely unknown.
In the present work, we estimated the prevalence of anti-Borrelia antibodies using a sample of 805 individuals belonging to nine seabird species breeding on six islands of the North Pacific. This large-scale sampling survey in the North Pacific was conducted to (1) investigate interspecific patterns of exposure to Bbsl in the North Pacific and compare these patterns with those found in the North Atlantic, (2) determine whether there is significant heterogeneity in the distribution of seropositive individuals among colonies within species, and (3) identify those seabird species or groups of species which may be important for the global circulation of Bbsl.
Materials and Methods
Detection of anti-Borrelia antibodies in seabirds
Blood samples were collected from 805 breeding adults belonging to nine seabird species: the crested auklet Aethia cristatella, the parakeet auklet Aethia psittacula, the least auklet Aethia pusilla, the common murre, the thick-billed murre, the tufted puffin Lunda cirrhata, the horned puffin Fratercula corniculata, the red-legged kittiwake, and the black-legged kittiwake. Sampling took place on six North Pacific islands (Fig. 1). In 2003 and 2004, samples were collected from Buldir (52°21′N 176°56′E), St. Paul (57°11′N 170°15′W), and St. George (56°35′N 169°37′W). In 2008, the west Pacific was sampled: Starichkov (52°46.919′N, 158°36.617′E), Ptichie (Ptichie 1 [50°30.495′N, 156°17.358′E] and Ptichie 2 [50°30.570′N, 156°16.762′E]), and the Commander Islands (Toporkov [55°12.414′N, 165°56.174′E] and Ary Kamen [55°12.842′N, 165°47.452′E]). Not all nine seabird species could be sampled on each island, notably because not all species are present at these locations (Hunt et al. 1986, Kondratyev et al. 2000, Byrd et al. 2005). We therefore aimed to sample the main species present on each island, so as to obtain representative samples of the local community. Sample sizes for each species and location are specified in Table 1. As sample sizes were small for the two locations of Ptichie and for the Commander Islands, we pooled the samples within each location. Blood samples from breeding adult birds were obtained from the left ulnar vein with a sterile syringe (Staszewski et al. 2008). After centrifugation, the plasma was separated from the blood cells and stored at −20°C until immunological analyses.
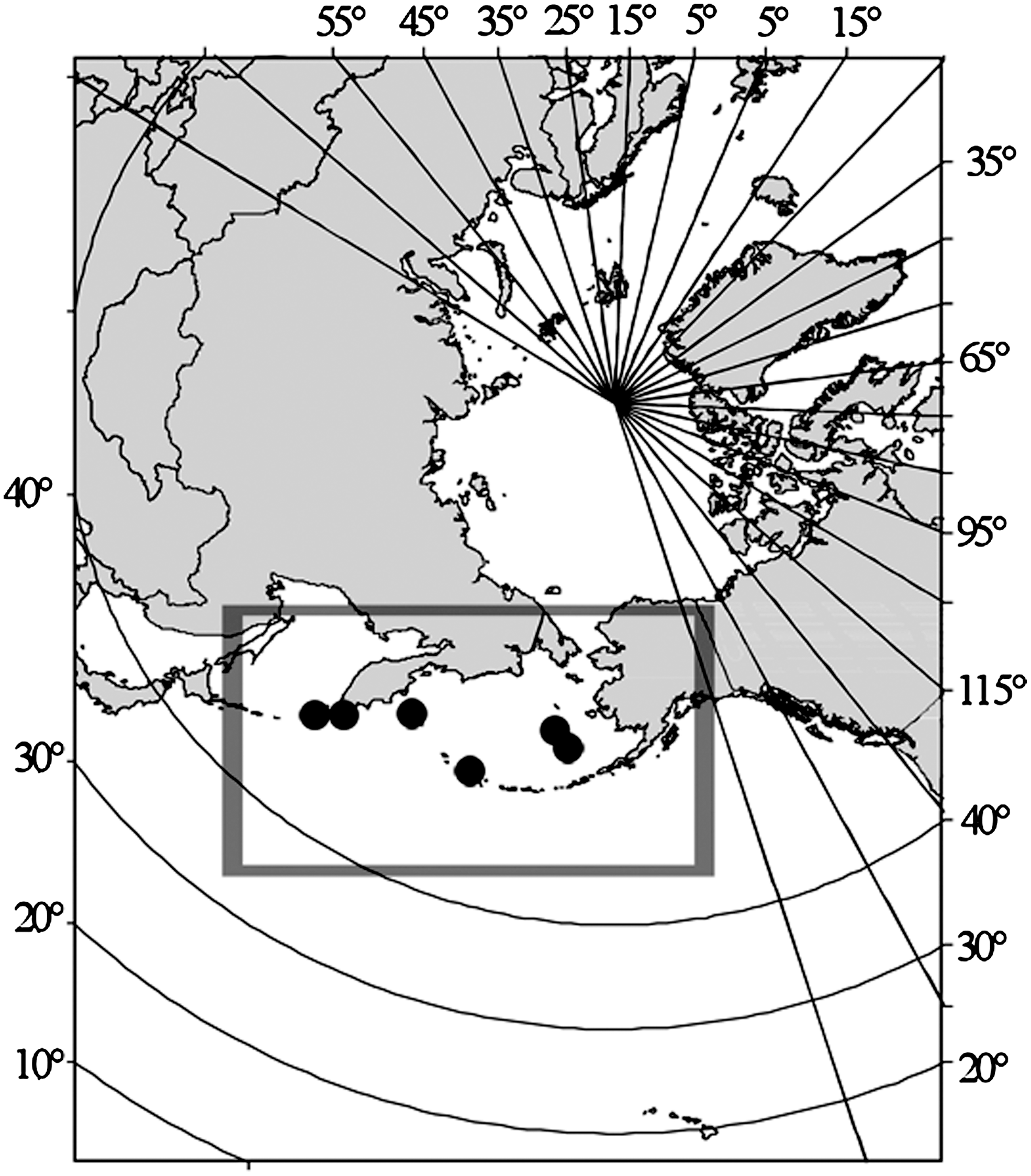
Geographic location of islands sampled in the North Pacific.
Antibody prevalence is expressed as the number of positive individuals divided by the total number of individuals analyzed. Numbers in square brackets express the standard deviation of prevalence calculated as sqrt (p *(1−p)). Fractions in brackets indicate the number of positive individuals with respect to the total number of individuals analyzed. Gray shading highlights the most abundant species in each large colony.
Anti-Borrelia Ab levels in the plasma were analyzed using a sandwich enzyme-linked immunosorbent assay (ELISA, Enzygnost Lyme VlsE/IgG, Dade Behring). Because this kit was manufactured for human use and was designed to recognize mammalian Ab, we replaced the anti-IgG Ab of the kit by an anti-chicken IgY (Sigma A-9171) conjugated with phosphatase. We diluted the samples in the buffer provided by the kit (dilution 1/50) and transferred 50 microliters to the ELISA plate wells. The plates were incubated 3 h at room temperature and then washed 3 times with phosphate-buffered saline (PBS)-Tween. Subsequently, we added 50 μL of anti-chicken Ab conjugated with phosphatase (dilution 1/2000) to the wells and incubated the plates overnight at 4°C. Plates were washed three times (PBS-Tween) and 100 μL of the phosphatase substrate p-nitrophenylphosphate-diethanolamine (1 mg/mL) was added to the wells. We measured the optical density at 405 nm at several points over time (every 2.5 min) to capture the dynamics of color development and saturation after the addition of the substrate chromogen. The optical density (OD) of the colorimetric reaction after 7.5 min was used for estimating the Ab levels in the samples as it gave us the best correlation between control samples run in two independent ELISA plates. A group of 8 samples representing a range of optical densities was run on each ELISA plate. A correction equation was then calculated between the OD of these samples on a given plate and that of a chosen reference plate; this equation (corrected OD=a×(original OD) + b) was then used to correct the OD of samples on a given plate (all regressions, r>0.96). Most samples were run once in a single ELISA well. However, intraplate coefficients of variation were calculated by running two to five samples more than once on each plate. Interplate coefficients of variation were calculated using the corrected OD of samples run on different plates.
To verify that immunoglobulins from the seabird species considered here were recognized by anti-chicken antibodies (Martinez et al. 2003), we carried out western blots. For this, plasma samples of the 9 seabird species were diluted 1/10 in SDS-reducing buffer (Laemmli sample buffer containing 0.5% mercaptoethanol). Plasma proteins (3 μL diluted 1/10) were separated on polyacrylamide gels containing a stacking gel of 4% and a running gel of 10%. Prestained markers of molecular weight (Spectra Multicolor Broad Range Protein Ladder; Fermentas Life Sciences) were included in all gels. Proteins were transferred to nitrocellulose membranes (Bio-Rad). Blots were then washed with PBS-Tween 0.05% and incubated overnight at 4°C with 5% nonfat dry milk (Regilait) in PBS-Tween. Membranes were then incubated for 2 h with an anti-chicken anti-IgY conjugated with peroxidase (Sigma, A-9046) diluted to 1/5000 in 3% nonfat dry milk in PBS-Tween. After 3 washes with PBS-Tween, antigen-Ab complexes were observed by chemoluminiscence using the SuperSignal West Pico reagent (Fisher). The anti-chicken Ab proteins detected corresponded to the apparent molecular weights of heavy and light immunoglobulin chains for all nine seabird species considered in this study (Fig. 2a).
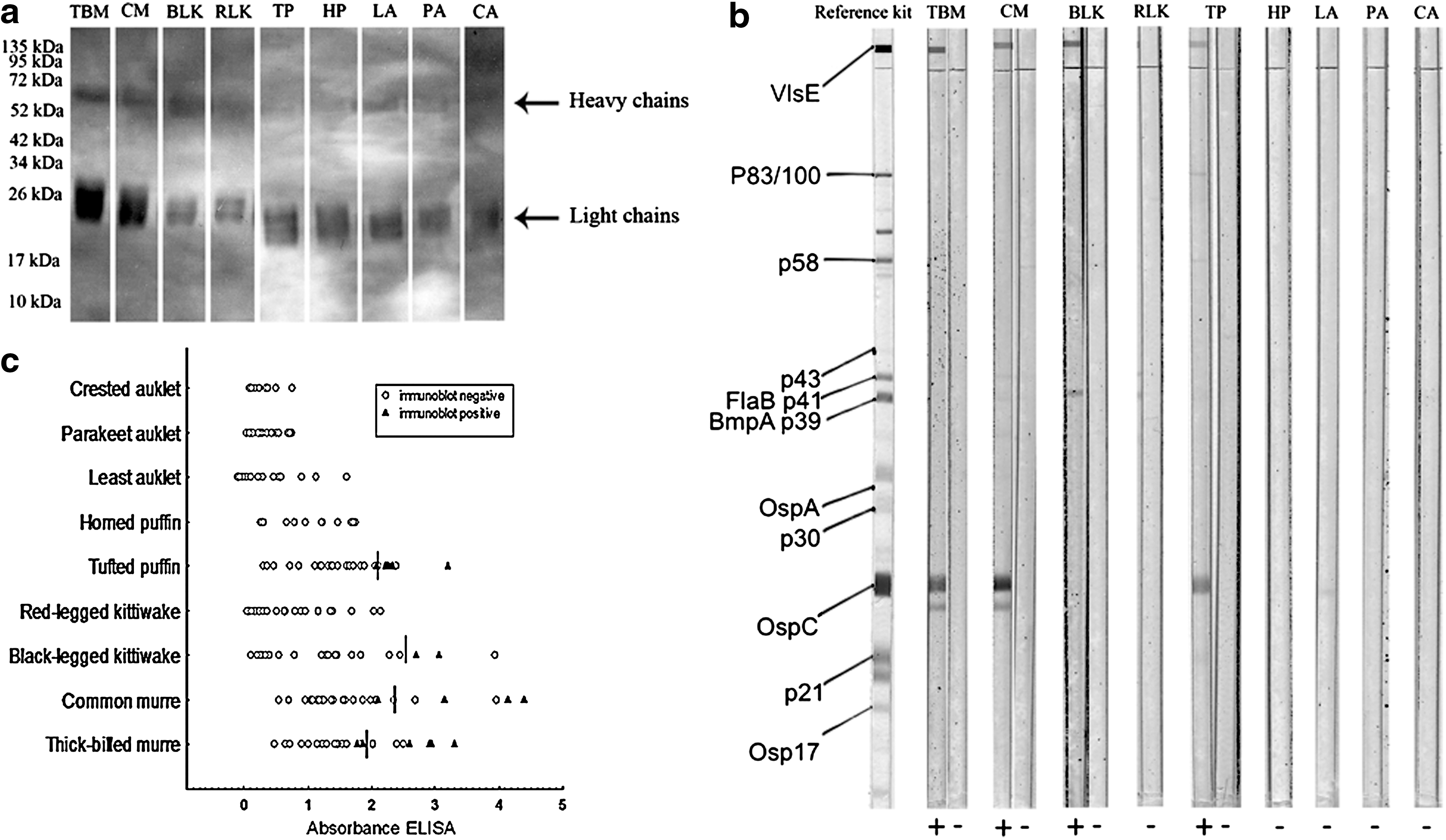
While the ELISA gave us a relative estimate of Ab levels, it was necessary to define specific positivity thresholds as the affinity of anti-chicken antibodies for IgY can vary among the species considered (Staszewski et al. 2008). We did this by performing western blots for the detection of antibodies against Bbsl (EcoBlot Borrelia afzelii+ VlsE; Meridian Bioscience) using a range of plasma samples covering the spectrum of optical densities for each species. Due to sample availability and to optimize costs, the number of samples tested in western blots analyses was 13 for crested auklets, 16 for parakeet auklets, 15 for least auklets, 12 for horned puffins, 27 for tufted puffins, 20 for red-legged kittiwakes, 24 for black-legged kittiwakes, 31 for common murres, and 23 for thick-billed murres. The antigens used in the kit come from the strain of Borrelia afzelii PKo and recombinant VlsE. The antigens relevant for diagnosis used by the kit are VlsE, p83/100, p58, p43, BmpA (p39), OspA (p31), p30, OspC (p23), p21, and Osp17; the presence of specific antibodies against each antigen is revealed by the presence of color bands at specific positions. The birds of our study were most likely exposed to Borrelia garinii, as suggested by the genetic studies of Borrelia genospecies isolated from ticks collected from some of these hosts and locations (Gómez-Díaz et al. 2011), although B. burgdorferi sensu stricto and B. afzelii have also been recorded in the marine system (Dietrich et al. 2008, Duneau et al. 2008). The proteins used in serodiagnosis are largely shared by the different species of the Bbsl complex, which should enable the detection of antibodies against these proteins from the different Bbsl species/strains. Moreover, the kit contains a recombinant VlsE, recommended to improve sensitivity to the presence of a variety of Borrelia strains (Wilske et al. 2007). We used the protocol suggested by the kit manufacturer, but substituted the anti-human IgG provided by the kit with anti-chicken IgY (Sigma A-9171) (Staszewski et al. 2008). Samples were diluted 1/100 and the anti-IgY was used at dilution 1/200. We measured the intensity of the bands using the program ImageJ and declared a band positive if its intensity was higher than the VlsE band of the positive control provided by the kit. A sample was declared positive if at least 2 positive bands were found (see Fig. 2b for examples of positive and negative immunoblots). This criterion probably gave us a conservative estimate of seropositivity. We ran 10 samples in 2 different immunoblots to calculate a Spearman correlation of band intensities of the different antigens (antigens positive in at least 3 samples: (BmpA (p39), OspA, OspC, p58, and VlsE) and summarized the results by calculating the average of the correlations.
The positive ELISA threshold of each species was calculated as the mean optical density minus one standard deviation of all individuals found positive by the western blot (Fig. 2c). An individual was declared positive for the ELISA if its optical density was above this threshold. This method has been used in previous studies on Borrelia seroprevalence in seabirds and with this calibration, ELISA results can be directly compared among individuals and species (Staszewski et al. 2008).
Differences in prevalence (percentage of seropositive individuals) among species and locations were analyzed using Fisher exact tests, testing each explanatory factor separately. To analyze the effect of species and locations simultaneously, we used only those locations where at least three seabird species were sampled and performed a logistic regression to evaluate the effect of the two explanatory variables on seroprevalence.
Results
For the ELISA tests, the average intraplate coefficient of variation was 6.85% (95% confidence interval [CI] 4.83–8.87, N coefficients of variation=39). The average interplate coefficients of variation was 30.04% (95% CI 27.01–33.07) for uncorrected values and 11.25% (95% CI 5.74–16.76, N coefficients of variation=24) after correction. For the western blot analyses, band intensities were highly correlated between replicate immunoblots (average Spearman r=0.86±0.17, N=5 antigens; BmpA (p39), OspA, OspC, p58, and VlsE).
The global prevalence of anti-Borrelia antibodies in adult seabirds was 7.0% (positives=56, N=805). No anti-Borrelia Ab was detected in five seabird species: the crested auklet (N=41), the parakeet auklet (N=75), the least auklet (N=96), the horned puffin (N=27), and the red-legged kittiwake (N=101). In the four other species in which anti-Borrelia antibodies were detected, prevalence differed among species (Fisher exact test p<0.001, N=465): common murre 18.1%, thick-billed murre 18.2%, tufted puffin 13.3%, black-legged kittiwake 2.5% (see Table 1 for details). There was a significant difference in the overall prevalence of Borrelia among islands (Fisher exact test p<0.001, N=805): Buldir 4.6%, St. Paul 3.6%, St. George 4.5%, Starichkov 7.5%, Ptichie 5.4%, and Commander 17.2%. When considering within species geographic variation in seroprevalence for those species with nonzero prevalence, significant differences in prevalence were observed among islands for common murres and black-legged kittiwakes (Fisher exact test p<0.05, Fig. 3): common murres showed the highest prevalence in Commander islands (58.6%) and black-legged kittiwakes showed the highest prevalence on St. George (16.7%). No significant island effect was observed for thick-billed murres and tufted puffins (Fisher exact test p>0.05; Table 1, Fig. 3). When considering only those locations where at least three species were sampled together (St. Paul, St. George, and the Commander Islands for common murres, thick-billed murres, and black-legged kittiwakes), both species and island effects were significantly associated with Ab prevalence (logistic regression, island effect: Wald=13.35, p=0.0013, species effect: Wald=12.74, p=0.0017, n=227).
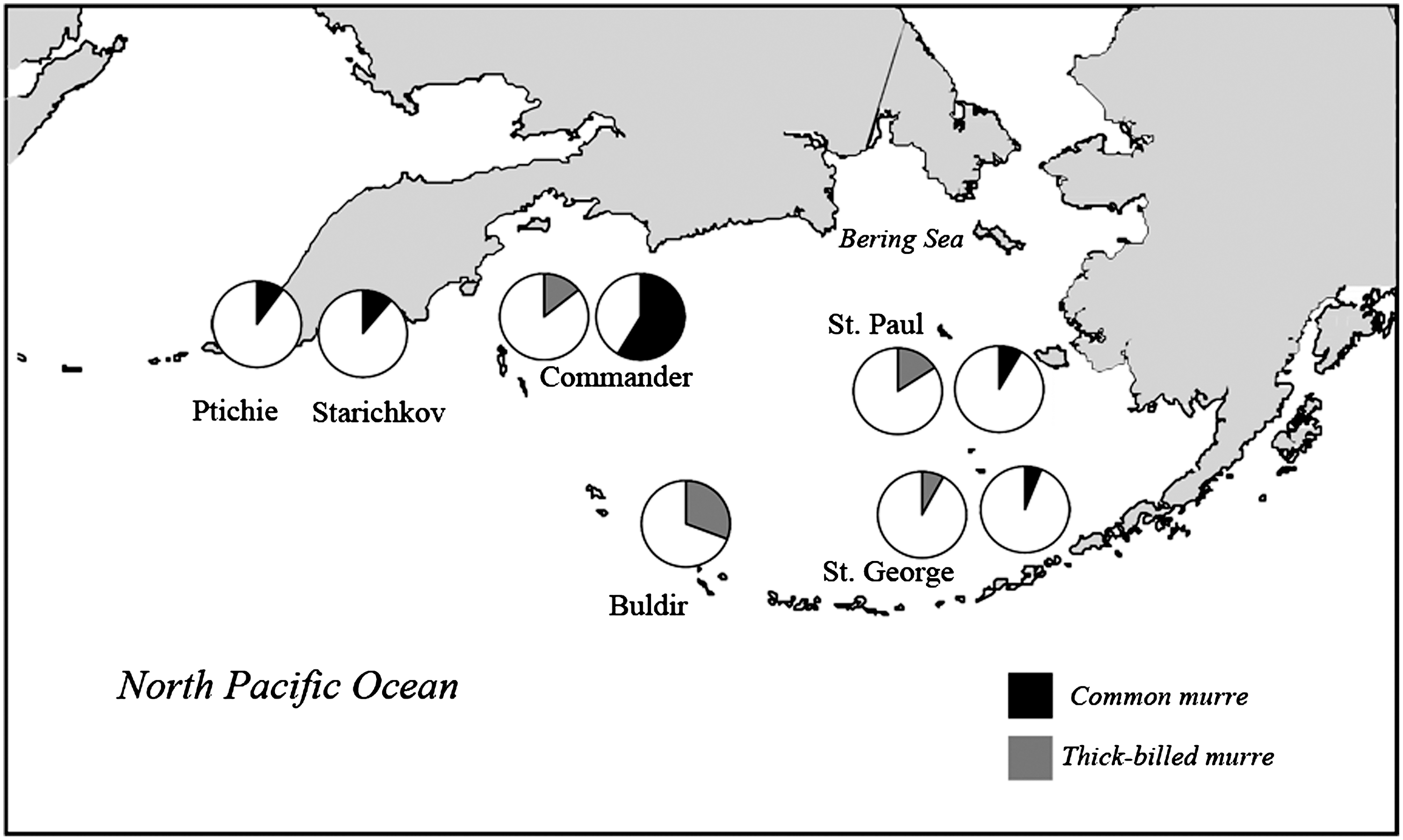
Prevalence of antibodies against Borrelia burgdorferi sensu lato in common and thick-billed murres. The painted area on each pie chart refers to seropositive individuals, and the white area refers to seronegative individuals for each species and location. Sample sizes are specified on Table 1. St. Paul, St. George, and Commander Islands show two graphs; one for each murre species.
Discussion
We demonstrate significant variability in Borrelia seroprevalence among seabird species in the North Pacific. Specific anti-Bbsl antibodies were measured by a quantitative ELISA technique using commercially available secondary anti-chicken antibodies. We defined a positive ELISA threshold for each seabird species using a qualitative western blot technique and verified that immunoglobulins of the studied seabird species were recognized by anti-chicken antibodies. These steps allowed us to determine Bbsl exposure in the nine focal seabird species. Only four of these nine species showed evidence of an immune response to Bbsl, suggesting that some species are unlikely to contribute to Borrelia circulation. Likewise, seropositive species differed in their relative prevalence; the highest seroprevalence was found for the two murre species, a result consistent with observations in the North Atlantic (Staszewski et al. 2008). Consequently, among the seabird species sampled, murres may be the main species implicated in the global circulation of Borrelia spirochetes and should be targeted for further study, especially with respect to the duration of infection, reservoir competence and dispersal.
Interspecific differences in Ab prevalence may be due to several alternative factors. First, there may be strong inter-host variability in exposure to tick vectors. For instance, high breeding densities in surface-nesting murres may increase their exposure to ticks compared with auklets, kittiwakes, and puffins, which usually breed at lower densities in individual burrows, nests or crevices (Cramp and Simmons 1983, Gaston and Jones, 1998). It is also possible that some species repel ectoparasites such as ticks by producing volatile odorants, as is the case in crested auklets (Douglas et al. 2004). However, differences in anti-Borrelia Ab prevalence cannot be explained purely by differences in vector presence. Some seabirds, such as red-legged kittiwakes and parakeet auklets, showed 0% prevalence of Bbsl antibodies, but are known to host I. uriae and to be present in the same colonies as seropositive birds (see Table 1). Alternatively, differences in seroprevalence may be due to host specialisation in tick vector. In the North Atlantic and sub-Antarctic, evidence of host-associated genetic structure has been found for I. uriae (McCoy et al. 2001, 2005b, Kempf et al. 2009), along with differences in Borrelia prevalence and intensity among sympatrically occurring tick races (Duneau et al. 2008, Gómez-Diaz et al. 2010). Information on the population genetic structure of I. uriae within North Pacific colonies will be now required to test the potential role of vector specialisation in determining patterns of Bbsl exposure. Finally, the different seabird species considered here may represent different environments for microparasites in terms of the resistance conferred by their immune system. Indeed, acquired resistance has been suggested to explain differential detection of Great Island virus in ticks feeding on juvenile and adult common murres (Nunn et al. 2006a, 2006b). The immune response has also been associated with observations of interspecific differences in reservoir competence of different host species of LB spirochetes in terrestrial systems (Kurtenbach et al. 2006).
Our results also indicated significant spatial variability in anti-Borrelia seroprevalence among locations. Anti-Borrelia Ab prevalence in birds was highest in the Commander Islands; this is consistent with a high spirochete prevalence reported in I. uriae ticks collected from the vegetation on these islands (Comstedt et al. 2009). The global and local prevalence of anti-Borrelia antibodies we observed in the North Pacific is, nevertheless, much lower than that observed in the North Atlantic for black-legged kittiwakes (global prevalence of 18.6%) and common murres (global prevalence of 77.1%) (Staszewski et al. 2008). The extent to which community composition explains differences in seroprevalence among Atlantic and Pacific colonies may strongly depend on within host species spatial variability in tick exposure (Gasparini et al. 2001, Staszewski et al. 2007) and the potential transmission constraints imposed by host specialisation in the tick vector (Duneau et al. 2008). Other environmental factors that differ between regions and affect food availability, host physiological states, and inter-colony movement patterns could also affect host infection status and pathogen prevalence (Gylfe et al. 2000, Kitaysky et al. 2001). To better understand the implications of interspecific and inter-locality differences in host prevalence in relation to parasite transmission, future studies will need to examine the relative contribution of host ecology, vector ecology, and immunology to the patterns observed here. A better identification of the links that exist between the marine and terrestrial cycles of Bbsl is also needed to understand large-scale eco-epidemiological patterns of human Lyme disease.
Footnotes
Acknowledgments
We thank John Citta, Chris Barger, Morgan Benowitz-Fredericks, Tom Dempsey, Ine Dorresteijn, Hector Douglas, Ann Harding, Anne Ploshnitsa, Mike Shultz, Rebecca Young, and Sarah Youngren, for their help during collection of samples in the field. Permissions to work in the study areas were obtained from local authorities. Sampling of birds in the eastern North Pacific was authorized by the Institutional Animal Care and Use, University of Alaska Fairbanks, and under all required federal and state permits. Dr. Nikolay N. Pavlov, Director, Komandorskiy State Nature Biosphere Reserve, Dr. Yuri Artukhin and Dr. Sergey V. Zagrebelny are commended for their professional guiding and logistic support. We thank Bruno Buatois (Plate-forme d'Analyse Chimique, CEFE) and Rudolphe Hamel and Dorothée Misse (MIVEGEC) for technical support. Thanks to Doris Gómez and Julien Vézilier for their help with figures. We thank two anonymous reviewers for their constructive comments. E. L. was supported by the program “Contratos para Investigación Posdoctorales, incluidas las ayudas para Becas Mec/Fullbright y Cátedras Príncipe de Asturias” from “Ministerio de Ciencia e Innovacion” of Spain. J. P.-D. was supported by a Chateaubriand postdoctoral fellowship from the French government. E. G.-D. was supported by a Marie Curie fellowship no. PIEF-GA-2008-221243. Field work and analyses were funded by grants from North Pacific Research Board, projects 320, B67, B77 to ASK and EPSCoR University of Alaska Fairbanks, CNRS Interdisciplinary Emerging Infectious Disease program, the French Polar Institute (Institut Polaire Français Paul Emile Victor program 333 on the ecology of dispersal of seabirds and ticks), and the Agence National de la Recherche (ANR-06-JCJC-0095-01)).
Disclosure Statement
No competing financial interests exist.