
Abstract
Rickettsia felis causes flea-borne spotted fever in humans worldwide. The cat flea, Ctenocephalides felis, serves as vector and reservoir host for this disease agent. To determine the role of dogs as potential reservoir hosts for spotted fever group rickettsiae, we screened blood from 100 pound dogs in Southeast Queensland by using a highly sensitive genus-specific PCR. Nine of the pound dogs were positive for rickettsial DNA and subsequent molecular sequencing confirmed amplification of R. felis. A high prevalence of R. felis in dogs in our study suggests that dogs may act as an important reservoir host for R. felis and as a potential source of human rickettsial infection.
Introduction
Rickettsia felis has been traditionally grouped as a member of the spotted fever group (SFG) of rickettsial organisms (Bouyer et al. 2001). Some researchers, however, have classified it as part of the “transitional” group because of the presence of phenotypic and genetic anomalies in R. felis, which include the presence of plasmids and its ability to serologically cross-react with both typhus group (TG) and SFG rickettsiae (Gillespie et al. 2007). R. felis was first detected in the cat flea, Ctenocephalides felis (Adams et al. 1990), and is an emerging human pathogen that causes flea-borne spotted fever or cat flea typhus throughout the world. The first case of flea-borne spotted fever in humans was reported in 1994 in Texas in the United States (Schriefer et al. 1994a). Since then, a number of human cases have been reported worldwide including Spain (Oteo et al. 2006), Germany (Richter et al. 2002), Mexico (Zavala-Velazquez et al. 2000), and Kenya (Richards et al. 2010). More recently, a cluster of five patients were diagnosed with R. felis infection in Victoria, Australia (Williams et al. 2010). Clinical signs of human flea-borne spotted fever include pyrexia, headache, malaise, myalgia, rash, and eschar (Richter et al. 2002, Richards et al. 2010). These signs are very similar to those caused by related rickettsial species (Schriefer et al. 1994a, Mert et al. 2006, Unsworth et al. 2007).
The first detection of R. felis was from a laboratory flea colony in the United States (Adams et al. 1990). Since then, the distribution of R. felis infection in different flea species collected from dogs and cats, as determined by molecular methods, has been found to be global, with infection rates of 15% in New Zealand (Kelly et al. 2004), 16% in Lebanon (Mba et al. 2011), 43.6% in Spain (Nogueras et al. 2010), 67.4% in the United States (Hawley et al. 2007), and 81% in New Caledonia (Mediannikov et al. 2011).
The cat flea, C. felis, has been reported to be both the primary vector and reservoir of R. felis (Reif and Macaluso 2009). Vertical transmission through transovarial and transstadial transmission of R. felis in the cat flea has been reported and plays an important role in the maintenance of this pathogen in the environment (Azad et al. 1992, Wedincamp and Foil 2002). Despite this progress in understanding the biology of R. felis, a definitive host has not been identified and clinical signs of infection have been reported only in humans. Nondomestic or wild animals including opossums and feral raccoons have been also shown to harbor R. felis (Schriefer et al. 1994b, Sashika et al. 2010). Although they have been implicated as potential mammal hosts, their roles as reservoir hosts for human infection requires further elucidation.
Although the domestic cat has been previously implicated as a potential primary reservoir for R. felis (Higgins et al. 1996, Case et al. 2006), recent evidence from a number of studies does not support this hypothesis. A prevalence study using molecular techniques reported 19.8% of flea sets collected from cats in eastern Australia harbored R. felis DNA (Barrs et al. 2010). However, the pathogen was not detected in the blood of these cats, and hence, it was speculated that domestic cats are unlikely to act as the primary vertebrate reservoir (Barrs et al. 2010). Studies conducted in the United States (Bayliss et al. 2009) and Canada (Kamrani et al. 2008) using gltA and/or ompB gene amplification on high-risk groups of cats did not result in detection of R. felis.
R. felis DNA has been detected using PCR assays in cats' blood in an experimental infection study (Wedincamp and Foil 2000) and in skin biopsy and gingival swabs of cats (Lappin and Hawley 2009) in the United States. However, natural infection in cats with active rickettsemia has not been verified by PCR assays. As a reported host for C. felis, dogs could potentially act as a reservoir for human flea-borne spotted fever. This is supported by a number of molecular studies, some carried out in Australia, which have demonstrated R. felis in a variety of fleas and ticks collected from dogs (Kelly et al. 2004, Schloderer et al. 2006). To date, there are only two reports outlining detection of R. felis DNA in dogs. In Spain, a PCR assay was used to detect R. felis in the blood of a dog owned by two people diagnosed with flea-borne spotted fever (Oteo et al. 2006). This dog was afebrile but showed signs of fatigue, vomiting, and diarrhea. A case report from Germany used western blotting to demonstrate R. felis infection in an asymptomatic dog owned by two patients with flea-borne spotted fever (Richter et al. 2002). Moreover, a seroprevalence study of R. felis in Spain reported that 51.1% of dogs were seropositive to this pathogen (Nogueras et al. 2009).
The present study thus aims to determine whether dogs may harbor R. felis by employing molecular techniques in a targeted prevalence survey on a high-risk group of poorly cared-for pound dogs from Southeast Queensland (SE QLD) in Australia.
Materials and Methods
Geographical area
The study was undertaken in SE QLD, Australia (Fig. 1). Brisbane, which represents the major metropolitan area of SE QLD, is the capital city of Queensland. It is situated in a subtropical region with warm, humid summers and short, mild winters.
The Southeast Queensland region of Australia, showing the location of the major metropolitan area, Brisbane.
Sampling
Samples were collected from January 2010 through to June 2010 from pound dogs sourced from the SE QLD area. Two EDTA-anticoagulated whole-blood samples were collected from 100 pound dogs by venipuncture in the Clinical Studies Centre, School of Veterinary Science, University of Queensland. Sex and estimated age were recorded prior to sample collection. One of the EDTA-anticoagulated whole-blood samples from each dog was sent to either IDEXX Laboratories in Brisbane or the veterinary clinical pathological laboratory at School of Veterinary Science, University of Queensland, for basic hematological evaluation, including a complete blood count. The remaining blood sample was stored at −20°C until DNA extraction.
DNA extraction
DNA was extracted from whole-blood samples collected from dogs into EDTA tubes. The DNeasy Blood & Tissue Kits (Qiagen) were used and the manufacturer's protocol followed with minor modifications. The blood volume used was increased to 200 μL and the final elution volume was decreased to 50 μL to yield more concentrated target DNA.
Molecular detection
DNA samples were tested using a previously described genus-specific PCR assay (Paris et al. 2008). The assay was performed using primers ompB-F 5′-CGACGTTAACGGTTTCTCATTCT-3′ and ompB-R 5′-ACCGGTTTCTTTGTAGTTTTCGTC-3′ targeting a 297-bp region of the outer membrane protein B (ompB) gene common to SFG rickettsiae. Two microliters of extracted DNA was added to a 23 μL reaction mixture containing 5× PCR buffer, 200 μmol/L dNTP, 1.0 mmol/L MgCl2, 0.5 units of GoTaq polymerase (Promega), 10 pmol of each forward and reverse primer, and a final volume of nuclease-free water. DNA from Rickettsia conorii israelensis (SFG) and nuclease-free water were used as positive and negative controls, respectively.
PCR amplification was performed with the initial activation step at 95° for 3 min, followed by 40 cycles of amplification at 95°C for 30 s, 54°C for 30 s, and 72°C for 30 s, and a final extension step of 72°C for 7 min. Amplified product was examined on 1.6% agarose gels stained with SYBR Safe (Invitrogen) run at 100 V for 30 min and visualized using UV transillumination. Positive PCR products were purified using the QIAquick PCR Purification Kit (Qiagen) according to manufacturer's protocol and submitted to the University of Queensland Animal Genetics Laboratory for DNA sequencing.
Phylogenetic analysis
DNA sequences were analyzed using Finch TV 1.4.0 (Geospiza, Inc.) and aligned using BioEdit version 7.0.5.3 (Hall 1999) with the ompB gene of the following rickettsiae species: R. felis, R. conorii, R. honei, R. asiatica, R. tasmanensis, R. rickettsii, and R. australis (GenBank accession no. GQ385243, AF123726, AF123711, DQ110870, GQ223393, X16353, and AF123709, respectively). Neighbor-joining analyses were conducted with Tamura-Nei parameter distance estimates, and trees were constructed using Mega 4.1 software (
Statistical analysis
The prevalence and 95% confidence intervals were calculated for PCR results using Win Episcope 2.0. The association between PCR results with host factor (age and gender), time of sample collection (summer, autumn, and winter), blood profile parameters including packed cell volume, erythrocyte count, hemoglobin, and leukocyte count was studied using chi-square test or Fisher's exact test for independence. Continuous data (blood parameters) were analyzed using one-way analysis of variance. Significance was set at p≤0.025. Univariate analyses were conducted using SPSS version 17.0 software (SPSS, Inc.) and Excel 2007 (Microsoft).
Animal ethics
This project was approved by the University of Queensland Animal Ethics Committee (Approval No.: SVS/419/09/BAYER/ARC LINKAGE).
Results
Of the 100 pound dogs, 56 were male and 44 female. Fifty-three blood samples were collected during summer (January and February), 32 during autumn (March–May), and 15 during winter (June). The sampled group was represented by a single puppy (<12 weeks), 24 juveniles (12 weeks–1 year), 68 adults (1–10 years), and 7 geriatrics (>10 years). Most dogs were of mixed breed. All dogs appeared healthy.
Nine (9%) of 100 pound dogs' blood were positive for Rickettsia by PCR. The sequences amplified from all nine PCR-positive dogs showed 99.7% similarity to the existing R. felis ompB sequence present in GenBank (Accession No. GQ385243). Phylogenetic analysis of a 297-bp region of the ompB gene (Fig. 2) closely grouped all nine sequences with R. felis GQ385243.
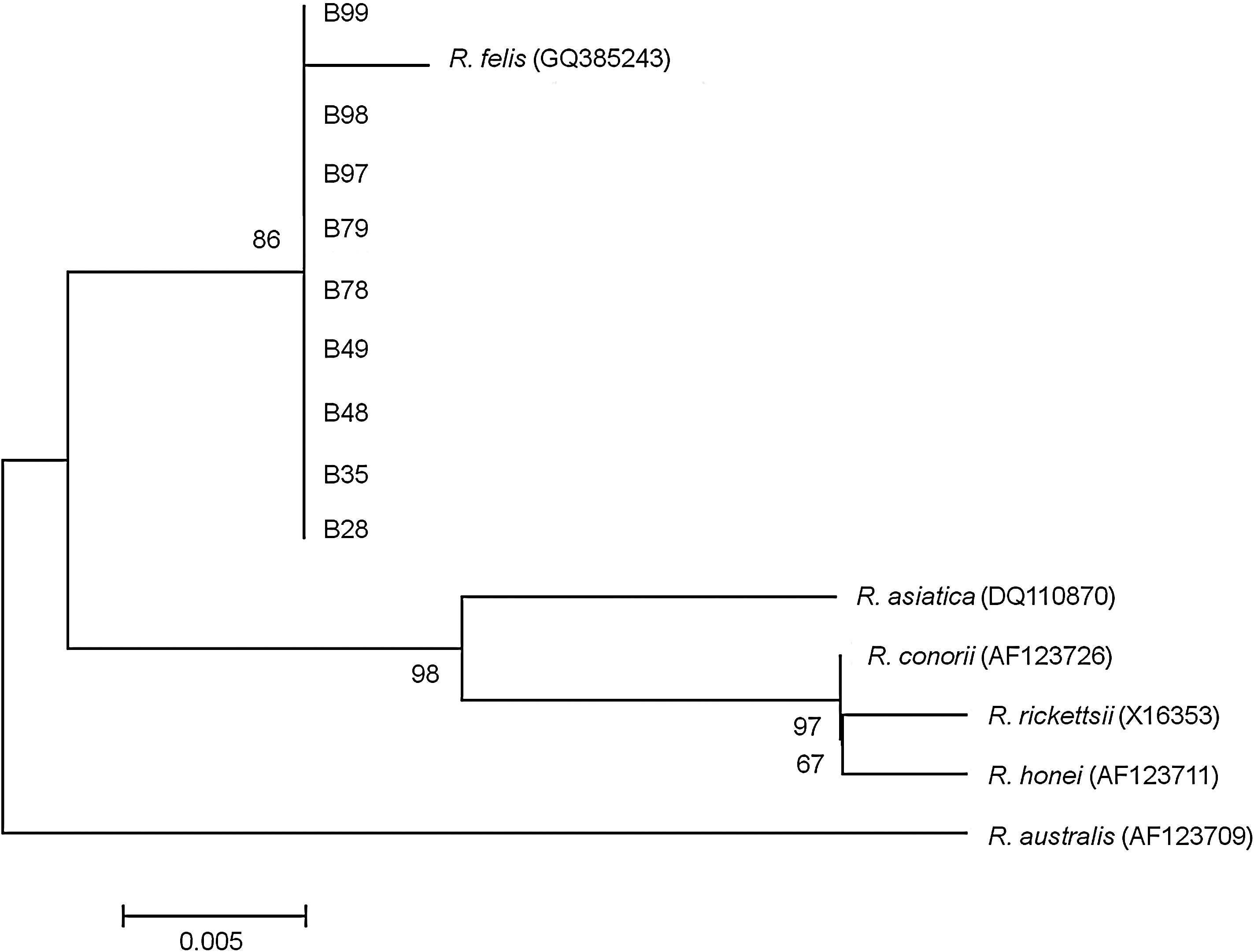
Phylogenetic tree obtained from neighbor-joining analysis of the ompB gene amplicons (297 bp) of Rickettsia sp. detected in nine pound dogs clustered with Rickettsia felis GQ385243.
Of these nine dogs, two were juveniles, six were adults, and one was geriatric. Seven dogs were male and two were female.
Blood profile parameters were obtained from all (n=97) except three dogs because of clotting of the blood sample. Two of the infected dogs were found to be mildly anemic (packed cell volume: 32%). However, 31 noninfected dogs were also found to be mildly anemic. There was no significant age or sex predisposition observed in PCR-positive dogs. There was also no significant difference in time of sample collection and blood profile parameters between infected and noninfected dogs.
Discussion
Our study represents the first report of R. felis infection in dogs in Australia. Using a PCR assay targeting the ompB gene, 9% of dogs in this study were found to carry R. felis DNA. All amplicons were 99.7% homologous to R. felis (GQ385243) with only one nucleotide difference, which may suggest degree of genetic variability in different geographical isolates.
Our report also describes the first cluster of canine rickettsemia caused by R. felis in the world and diagnosed by PCR. Since the first recognition of R. felis in C. felis in 1990, household pets have been implicated as a potential reservoir as they stay in close proximity to humans and are frequently infested with C. felis. A high prevalence of R. felis in dogs in our study has shown that dogs may have the potential to act as an important reservoir and sentinel host for human infection (Richter et al. 2002, Oteo et al. 2006). Despite limited available data, multiple studies have been unsuccessful in detecting rickettsemia in cats (Hawley et al. 2007, Bayliss et al. 2009, Assarasakorn et al. 2010, Barrs et al. 2010), whereas detection of PCR positivity in dogs has been previously reported in Spain (Oteo et al. 2006). Dogs are also known to be ubiquitous, easily accessible, and susceptible to a wide range of emerging human diseases (Cleaveland et al. 2006).
To our knowledge, this study also describes the presence of Rickettsia spp. DNA in dogs' blood by PCR for the first time in Australia. Previous surveys of spotted fever rickettsial diseases in Australia involving dogs were conducted using serological assays (Sexton et al. 1991, Izzard et al. 2010). Rickettsial infection is difficult to specifically determine using serological tests alone because of cross-reaction among SFG Rickettsia spp. (Sexton et al. 1991). Moreover, the early classification of R. felis as part of the TG is because of its serotypical similarity to the TG rather than the SFG. Only when the presence of the ompA gene in the R. felis genome was demonstrated, it was reclassified into the SFG (Bouyer et al. 2001). Thus, diagnosis of R. felis infection or exposure via serological methods can be challenging and would reinforce the hypothesis that Rickettsia typhi cases in the past may have been misdiagnosed R. felis infections.
The pathogenicity of R. felis infection in dogs is unclear at this time. All the pound dogs in this study appeared healthy. To date, association of clinical disease and R. felis infection in animals has not been reported. Previously, in Spain, R. felis was detected by PCR in two patients and their dog, which showed signs of fatigue, vomiting, and diarrhea (Oteo et al. 2006). The authors did not elaborate on the clinical signs in this dog and no further work-up was performed.
Australia is endemic with rickettsial diseases, which includes epidemic typhus (Rickettsia prowazekii), murine typhus (R. typhi), scrub typhus (Orientia tsutsugumushi), Q fever (Coxiella burnetii), Queensland tick typhus (R. australis), and Flinders Island spotted fever (R. honei). This report, as well as three other studies on R. felis (Schloderer et al. 2006, Barrs et al. 2010, Williams et al. 2010), suggests that this pathogen is endemic in Australia and causes human infections. Following the detection of R. felis in this region, local health authorities should be aware of the occurrence of this flea-borne spotted fever and its zoonotic potential.
Flea infestation in dogs was not evaluated in the present study and a history of flea infestation in all these dogs could not be confirmed. Nevertheless, flea infestation was observed in some dogs. Fleas that are commonly found to infest dogs in Australia are C. felis (Schloderer et al. 2006), which is a nonhost specific flea. Other fleas that infest dogs in Australia are Ctenocephalides canis and Echidnophaga gallinacea (Schloderer et al. 2006), albeit rarely. R. felis infection in fleas collected from dogs and cats in Australia was first reported in 2006 (Schloderer et al. 2006). Recently, R. felis DNA was detected by PCR in 19.8% of flea sets collected from cats in eastern Australia (Barrs et al. 2010). In the present study, fleas were not collected, and thus the role of fleas as a vector for R. felis infection in dogs in SE QLD remains unclear. Future study on prevalence of R. felis in fleas and ticks sourced from dogs in this region is required to evaluate the epidemiology of this flea-borne spotted fever.
R. felis DNA has been detected in a variety of arthropods, including fleas, ticks, and mites, collected from a variety of mammals (Schloderer et al. 2006, Choi et al. 2007, Oliveira et al. 2008). Although ticks such as Rhipicephalus sanguineus (Oliveira et al. 2008), which infest dogs, were reportedly carrying this agent, it is possible that these arthropods may just feed on rickettsemic blood and act as mechanical or noncompetent vectors (Reif and Macaluso 2009).
Horizontal transmission between the cat flea and vertebrates remains poorly understood. Observation of R. felis in the salivary gland of C. felis under transmission electron microscope (Macaluso et al. 2008) suggests the potential for infection of vertebrates through blood feeding. Seroconversion in cats experimentally infected with R. felis through blood feeding by infected cat fleas (Wedincamp and Foil 2000) also suggests that horizontal transmission is possible. However, in this experimental infection study, rickettsemia in cats was found to subside rapidly. Horizontal transmission through blood feeding on infected cats and artificially infected meals, cofeeding between infected and uninfected fleas, and larval feeding on flea feces and eggs was not detected in a study by Wedincamp and Foil (2002). Hence, the transmission dynamics of R. felis infection in dogs remains an important area for future exploration.
In conclusion, detection of R. felis infection in dogs in this study is highly suggestive that dogs may act as a reservoir host for R. felis. However, this study was undertaken on a targeted high-risk group of dogs and was therefore not representative of a comprehensive prevalence survey. To better understand the natural epidemiology of R. felis in dogs, a more extensive survey is required, encompassing both high- and low-risk groups. Further studies are also required to study the transmission dynamics and pathogenicity of R. felis in dogs.
Footnotes
Acknowledgments
This study was supported by grants from Bayer Animal Health Australia and the Centre for Companion Animal Health, School of Veterinary Science, The University of Queensland.
Disclaimers
The opinions expressed by authors contributing to this journal do not necessarily reflect the opinions of the institutions with which the authors are affiliated.
Disclosure Statement
No competing financial interests exist.