
Abstract
We used epidemiological data and indirect fluorescent antibody tests to determine the Hantaan virus (HTNV) antibody-positive (Ab+) prevalence in small mammals captured at firing point 10 (FP-10) and firing point 60 (FP-60), Gyeonggi Province, near the demilitarized zone, Republic of Korea (ROK), from 2001 to 2005. We used these data, combined with the partial M segment amplified from HTNV recovered from lung tissues of Apodemus agrarius, to clarify the genetic diversity and phylogenetic relationships among HTNV strains in the ROK. Of the eight species of rodents and one insectivore species captured, A. agrarius accounted for 93.4% and 88.5% at FP-10 and FP-60, respectively. Only two species of rodents, A. agrarius and Micromys minutus, were HTNV Ab+. The overall HTNV Ab+ prevalence for A. agrarius captured at FP-10 and FP-60 was 23.3% (121/520) and 14.5% (94/647), respectively. The hantaviral reverse transcription–polymerase chain reaction-positive rate of Ab+ A. agrarius was 74.2% (167/215), and the phylogenetic trees, based on the 269-nucleotide G2-encoding M segment, demonstrated that HTNV strains from FP-10 and FP-60 were distantly segregated from HTNV of other geographic regions in Korea and China. These data are useful in the development of risk reduction strategies for the prevention of hantavirus infections among military personnel, especially during training or the event of hostilities, and civilian populations.
Introduction
Contained herein are epidemiological data that identify HTNV Ab+ rates for small mammal populations, characterization of HTNV strains, and biological and environmental conditions conducive for hantavirus transmission at firing point 10 (FP-10) and firing point 60 (FP-60). These data are useful for developing risk reduction strategies for preventing hantavirus infections in U.S. and ROK military personnel while training in field environments.
Materials and Methods
Comprehensive seasonal small mammal surveys were conducted in May and June 2001 at FP-10 and FP-60, respectively, through December 2005 as described by O'Guinn et al. (2008). FP-10 and FP-60 are ROK-operated training sites located in Yeoncheon-gun (county), Gyeonggi Province, near the DMZ (Fig. 1). Surveillance site selection was based on several factors: (1) previous reports of HFRS patients associated with exposure at FP-10 and FP-60, (2) habitats associated with relatively high rodent populations, and (3) sites that were adjacent to, but did not interfere with, military training activities.

Satellite photographs of FP-10
Small mammal habitat ecology
The primary small-mammal habitats at both firing points consisted of heavy to sparsely vegetated areas along the margin of artillery firing positions, cantonment, command and control, and maneuver sites that supported high small-mammal populations as described by O'Guinn et al. (2008). The central training area of FP-10 was largely barren, while FP-60 had irregularly distributed islands (10–20 m in diameter) of tall grasses, separated by 10–20 m of ground devoid of vegetation, which allowed for the dispersal of small mammals throughout the interior (Fig. 1). Habitats along some trap lines were modified due to construction, farming, or military activities, which resulted in more extreme seasonal vegetation changes and increased the potential movement and natural predation of small mammals and competition for more favorable habitat during the onset of the winter season.
Rodent trapping
Collapsible live-capture traps, Sherman® traps (7.7×9×23 cm; H.B. Sherman, Tallahassee, FL), were set at 4–5 m intervals (20–40/trap line) during daylight hours over 2 days as described by O'Guinn et al. (2008). Traps positive for small mammals were collected the following morning, sequentially numbered, and transported to Korea University in accordance with standard procedures for collection and transportation of specimens to minimize hazards from potentially infected rodents as described by Mills et al. (1995a, 1995b). The small mammals were anesthetized and euthanized by cardiac puncture under a Korea University animal use protocol, identified, sexed, weighed, and tissues (spleen, lung, and kidney) removed and stored at −70°C until used. All personnel processing rodents in the laboratory were vaccinated with a Korean-approved hantavirus vaccine (Hantavax®) with no laboratory cases reported.
Hantavirus serology and RT-PCR
Small mammal sera were diluted 1:16 in phosphate-buffered saline and examined for IgG antibodies against HTNV, Seoul, and Puumala viruses by indirect immunofluorescent antibody test (IFAT) (Lee et al. 1978, Song et al. 2007). Tissues from hantavirus Ab+ rodents (from sera) were initially assayed for antigens against hantaviruses by IFAT. Subsequently, lung tissues of all HTNV Ab+ rodents were used for the amplification of the HTN viral gene by RT-PCR. Total hantavirus RNA, extracted from lung tissues of Ab+ rodents and/or infected Vero cells using RNA-Bee (TEL-TEST, Inc., Friendswood, TX), was amplified by RT-PCR and reverse transcribed using the superscript II RNase H− reverse transcriptase kit (GIBCOBRL, Gaithersburg, MD). Primers (outer primer set, 5′-TGGGCTGCAAGTGC-3′ and 5′-ACATGC TGTACAGCCTGTGCC-3′; inner primer set, 5′-TGGGCTGCAAGTGCATCAGAG-3′ and 5′-ATGGATTACAACCCCAGCTCG-3′) afforded amplification of a 373-nucleotide (nt) region of the hantavirus G2 encoding M segment (Xiao et al. 1992, 1994, Song et al. 2000). Amplified products were size fractionated by electrophoresis on 1.5% agarose gels containing ethidium bromide (0.5 mg/mL). PCR products were cloned using the TA cloning system (Invitrogen Corp., San Diego, CA), and plasmid DNA was purified by the QIA prep-spin Plasmid kit (QIAGEN, Inc., Chatsworth, CA). DNA sequencing was performed in both directions from at least three clones of each PCR product, using the dye primer cycle sequencing ready reaction kit (Applied Biosystems, Foster City, CA) on an automated sequencer (model 3730; Applied Biosystems, Foster City, CA).
Genetic and phylogenetic analyses
Alignment and comparison of partial M segment sequences of four HTNV strains amplified from Apodemus agrarius (striped field mouse) from FP-10 and FP-60 with previously published hantavirus sequences were facilitated using the Clustal W method (Lasergene program version 5; DNASTAR, Inc., Madison, WI). For phylogenetic analysis, the neighbor-joining (N-J) and maximum parsimony (Phylogenetic Analysis Using Parsimony [PAUP] version 4.0b) methods and the unweighted pair-group method of assortment were employed. Genetic distances were computed by PAUP, and topologies were evaluated by bootstrap analysis of 1000 iterations (Swofford, 2003).
Results
Sampling
A total of 1288 small mammals (rodents and insectivores) were collected from FP-10 (557) and FP-60 (731), over 5700 trap nights (FP-10, 2620; FP-60, 3080), with an overall capture rate for all trapping periods of 20.6%. Seasonal capture rates for A. agrarius at FP-10 and FP-60 were significantly different (χ 2=23.8, df=3, p<0.001) and were likely affected by differences in seasonal reproductive rates and environmental conditions, for example, seasonal temperatures, cloud cover, and precipitation during each survey (Fig. 2). Overall, A. agrarius accounted for 90.6% (1167) of all the small mammals captured, followed by Crocidura lasiura (Ussuri white-toothed shrew and primary reservoir for Imjin virus, 6.2%, 80), Rattus norvegicus (Norway rat, 1.2%, 15), Mus musculus (house mouse, 0.7%, 9), Tscherskia (= Cricetulus) triton (greater long-tailed hamster, 0.5%, 6), Micromys minutus (harvest mouse, 0.4%, 5), Microtus fortis (reed vole, 0.2%, 3), Myodes (= Eothenomys) regulus (Royal vole, 0.2%, 2), and Rattus rattus (roof rat, 0.1%, 1). During 2003–2005, lines 1a and 1b at FP-10 were similarly trapped, and overall annual capture rates for the disturbed sites (line 1b), which had reduced ground cover (especially during the winter and spring season), were significantly lower (15.4%) (χ 2=65.4, df=4, p<0.001) when compared to the less disturbed area (line 1a) (24.3%). Conversely, the overall HTNV Ab+ rates were significantly higher for the disturbed habitat (31.0%) (χ 2=6.8, df=1, p=0.009) when compared to the relatively undisturbed habitat (15.3%). Although the HTNV Ab+ rates were lower for the undisturbed habitat, the estimated numbers of HTNV Ab+ mice collected/100 traps were nearly equal for the undisturbed and disturbed habitats (3.2 and 3.7, respectively) at FP-10. While only limited data were available for the more disturbed 10–20-m-diameter tall grass islands sampled during the winter of 2005 at FP-60, capture rates were significantly lower (23.3%) than the perimeter trap lines combined (39.4%) (χ 2=602.0, df=4, p<0.001) during the same period. However, HTNV Ab+rates (28.6%) were significantly higher for the tall grass islands than the less disturbed perimeter areas (17.5%) (χ 2=304.0, df=4, p<0.001) trapped during that same period, with estimated numbers of HTNV Ab+ mice/100 traps similar for the tall grass islands (6.7) compared to the perimeter habitats (6.9).

Capture rates for Apodemus agrarius by season (Sp, March–May; Su, June; F, August–September; W, November–December) for FP-10 and FP-60, 2001–2005. Sp, spring; Su, summer; F, fall; W, winter.
Overall capture rates for male and female A. agrarius were not significantly different (χ 2=0.7, df=1, p=0.410), with males accounting for 50.6% (263♂:257♀) and 53.0% (343♂:304♀) of those captured at FP-10 and FP-60, respectively. The overall mean percent of gravid A. agrarius females (2003–2005) was significantly higher during the fall trapping period (27.3%, range 24.5–32.6) (χ 2=196.9, df=3, p<0.001), while only 4.2% (range 3.7–6.1%), 1.3% (range 0–3.5%), and 0.3% (range 0–0.8) were gravid during the spring, summer, and winter trapping periods, respectively.
Serology studies
IgG antibodies against HTNV were detected by IFAT in 18.4% (215/1167) of A. agrarius. The only other seropositive rodent for HTNV was one of five (20%) M. minutus. That specimen was captured during the summer trapping period of 2003 at FP-60 when the seasonal Ab+ rate for A. agrarius was 12.2%. None of the other small mammal species tested was Ab+ for HTNV. Seasonal A. agrarius Ab+ rates ranged from 0% to 69.2% and 0% to 33.3% at FP-10 and FP-60, respectively, and were asynchronous, such that Ab+ rates were high during some trapping periods at FP-10 and low at FP-60 and vice-versa (Fig. 3). Annual Ab+ rates during 2001–2003 were higher at FP-10, but during 2004–2005 they were higher at FP-60 (Table 1). While the annual percent of Ab+ A. agrarius captured at both FP-10 and FP-60 were higher for males (mean 23.7%) than for females (mean 14.2%), they were not significantly different (χ 2=2.9, df=1, p=0.086). The overall percent of HTNV Ab+ A. agrarius captured at both FP-10 and FP-60, while variable seasonally, were similar for summer (16.6%) and winter (14.2%) surveys, but were significantly higher during the spring (21.9%) and fall (25.1%) trapping periods (χ 2=11.3, df=3, p=0.010) (Fig. 3). A. agrarius were weighed and placed in weight categories of 10-g intervals. Only a few A. agrarius that weighed <10 g were captured. Ab+ rates among the different weight classes increased similarly by weight class at both FP-10 and FP-60 (Table 2).
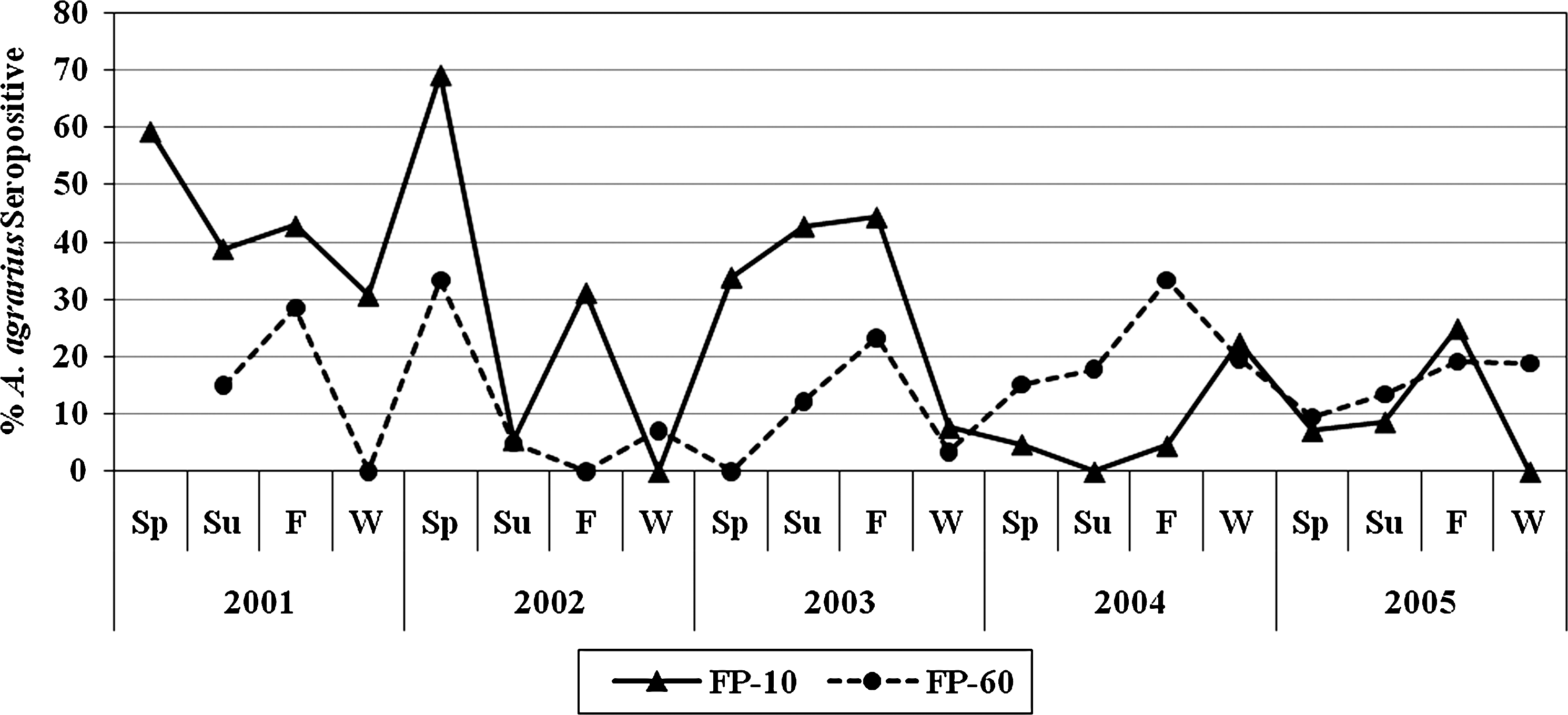
Seasonal HTNV antibody-positive rates for A. agrarius captured at FP-10 and FP-60 (Sp, March–May; Su, June; F, August–September; W, November–December), 2001–2005. In addition, one Micromys minutus was HTNV antibody positive during the summer trapping period of 2003 at FP-60. HTNV, Hantaan virus.
Percent (number) of Hantaan virus antibody positive (number tested).
FP-10, firing point 10; FP-60, firing point 60.
AB+, antibody positive.
Sequence analysis
Viral genomes of HTNV strains were amplified from tissues of 44 A. agrarius. The 269-nt G2-encoding M segment sequences (coordinates 2018 to 2298) of 20 HTNV strains from FP-10 (01-425, 01-431, 01-432, 01-443, 01-446, 01-447, 01-542, 01-577, 01-583, 02-185, 04-1257, 05-8, 05-325, 05-331, 05-344, 05-771, 05-775, 05-784, 05-810, and 05-1156) and 24 strains from FP-60 (01-429, 04-182, 04-187, 04-205, 04-211, 04-602, 04-604, 04-618, 04-630, 04-633, 04-938, 04-939, 04-944, 04-946, 04-948, 04-967, 04-1282, 04-1293, 04-1325, 04-1332, 05-309, 05-1057, 05-1227, and 05-1228) were sequenced. The GenBank numbers of 11 submitted HTNV strains are HM172504–HM172514. The nucleotide and amino acid divergence of the 20 HTNV strains from FP-10 were 0%–2.9% and 0%–2.2%, respectively. Those of the 24 HTNV strains from FP-60 were 0%–1.7% and 0%–1.1%, respectively. The nucleotide and amino acid differences between HTNV strains from FP-10 and FP60 were 1.1%–3.0% and 0%–2.2%.
The genetic distance of HTNV strains from the ROK and the People's Republic of China based on a 269-nt region of the G2 glycoprotein-encoding M segment was 9.6% to 12.5% at the nucleotide level.
Phylogenetic analysis
An N-J phylogenetic tree, based on the 269-nt region of the G2 glycoprotein-encoding M segment of HTNV, demonstrated that virus strains from A. agrarius from various HFRS-endemic areas in South Korea formed a monophyletic group, which was evolutionarily distinct from the Bao14 HTNV strain of the People's Republic of China (Fig. 4). Trees constructed by the N-J, maximum parsimony methods, and by UPGMA indicated that HTNV strains from FP-10 and FP-60 segregated well by their geographical origin except a few HTNV strains, including 05-1156FP10, 04-1282FP60, 04-1325FP60, and 04-602FP60. HTNV isolates in the ROK formed two distinct subgroups, with the prototype strain, 76–118, belonging to a subgroup distinct from those isolated from rodents captured at FP-10 or FP-60.
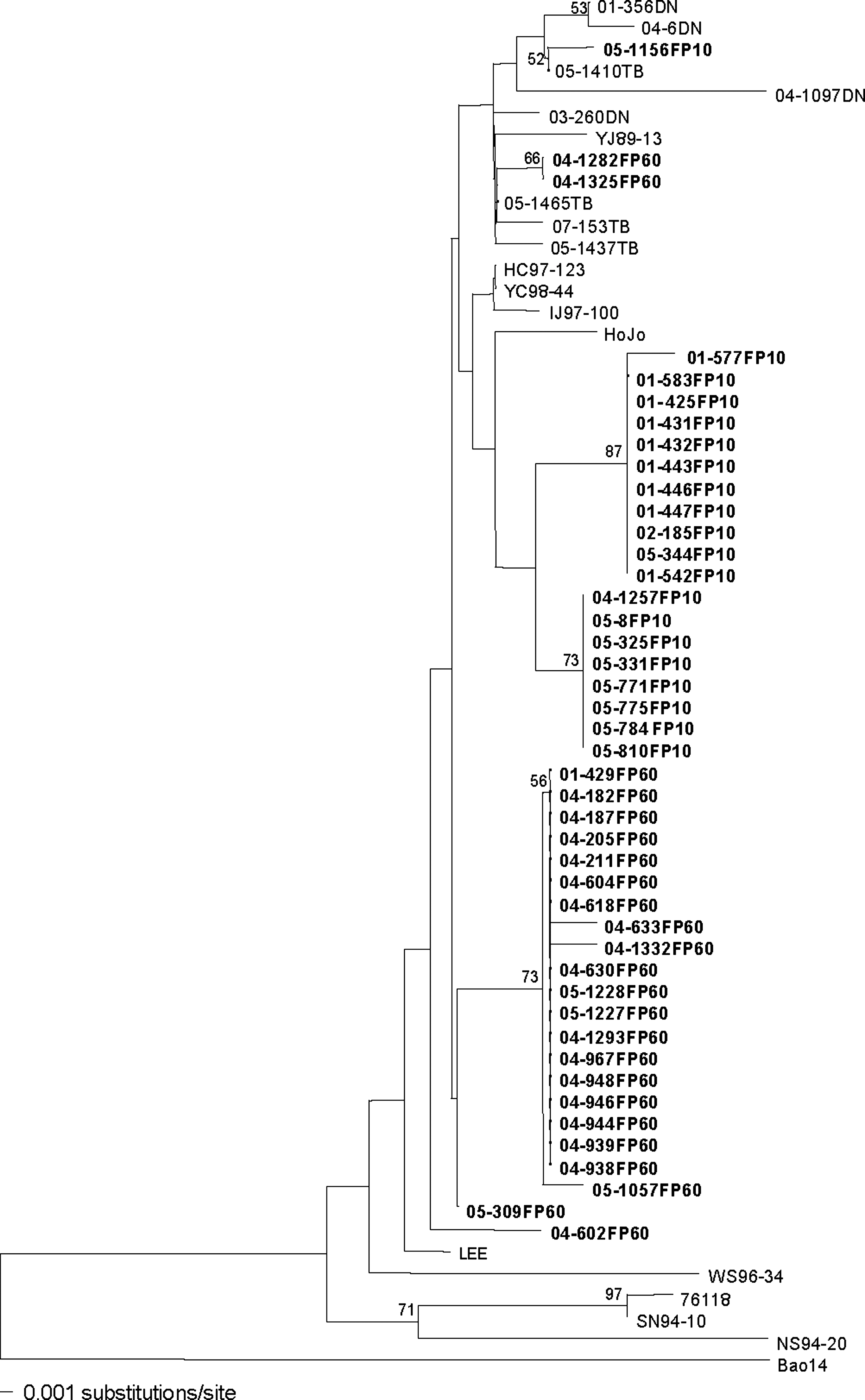
Neighbor-joining phylogenetic tree, based on the 269-nucleotide region of the G2 glycoprotein-encoding M segment of HTNV strains, from A. agrarius captured at FP-10 and FP-60 (bold character), Yeoncheon-gun, Gyeonggi Province, and HTNV strains at Dagmar North (DN), Twin Bridge (TB), Yangju (YJ), Dongducheon (prototype 76118 and SN) in Gyeonggi Province, Hongcheon (HC), Inje (IJ) in Gangwon Province, Nonsan (NS), Chungnam Province and Whasun (WS) in Jeonnam Province, Republic of Korea, showed phylogenetic distinct clustering of Korean and Chinese (Bao 14) strains. Branch lengths are proportional to the number of nucleotide substitutions, while vertical distances are for clarity only. The numbers at each node are bootstrap probabilities (expressed as percentages), as determined for 1000 iterations by PAUP version 4.0b.
Discussion
Rodent-borne disease surveillance is essential to identify the relationships between HTNV Ab+ rates, relative abundance of rodents, acute versus chronic infections, and associated human activities. This requires relatively large sample sizes of all ages (weights) to develop inferences for developing risk analysis. While few A. agrarius weighing ≤10 g were collected, which may be a result of trap sensitivity, C. lasiura weighing 5–10 g were captured in relative large numbers. In addition, only ∼1%–5% of the unsnapped traps had evidence (i.e., feces and/or removal or partially eaten bait) of small mammals in the traps. The current traps were used, as the use of smaller and perhaps more sensitive traps, would likely have reduced the capture rates of larger A. agrarius and other larger species, for example, M. fortis, T. triton, M. regulus, and Rattus spp. Overall, A. agrarius was the most commonly collected rodent in field environments with trap rates occasionally exceeding 50%. It is often found associated with tall grass habitats, especially along banks, with varying degrees of crawling and other herbaceous plants and shrubs associated with unmanaged lands and training sites (Bae et al. 1971, Song et al. 1971, Lee and Kim 1972, O'Guinn et al. 2008, Sames et al. 2009, Kim et al. 2011). However, during the winter (late November/December) and early spring (March), traps rates were generally lower where herbaceous plants, other than grasses and crawling vegetation that provided ground cover, predominated (i.e., line 1b, 2a, and parts of line 2c).
The relationships between (among) HTNV Ab+ rates, the relative density of striped field mice, environmental conditions, and incidence of human infection are complex and poorly understood. Fluctuations of rodent abundance and HTNV Ab+ rates have been linked to disease outbreaks, and the phenomena of increased viral shedding in acute infections likely impact on the potential for disease transmission during training operations (Mills and Childs 1998, Heyman et al. 2001, Olsson et al. 2002, 2003, 2005, Rose et al. 2003, Davis et al. 2005, Hardestam et al. 2008). However, unlike the bank vole, Myodes (= Clethrionomys) glareolus, where annual populations rapidly increase over a period of several years followed by sharp population declines that are positively correlated with numbers of HFRS cases, annual populations of A. agrarius and the number of HFRS cases (range 323–427 for 2001–2005) are relatively stable in the ROK (Lee et al. 1981, O'Guinn et al. 2008, Kim et al. 2011).
We found that HTNV Ab+ rates were positively correlated with weight (age) of A. agrarius. This is similar to other studies and is highly suggestive that the primary source of infection is horizontal, resulting from wounding or indirect transmission from HTNV in the environment, rather than vertical (transplacental or during nesting) (Glass et al. 1988, Douglas et al. 2001, Sauvage et al. 2002, Hinson et al.2004, Kallio et al. 2006, Sames et al. 2009; T.A.K., unpublished data). During the spring (March–April, 4.2%–40%) and the fall (August–September, 24.5%–70%) periods at various training sites, there were significant increases in the proportions of gravid females (O'Guinn et al. 2008; T.A.K., unpublished data). During the spring, when few HFRS cases are observed, there is an expansion of new growth of herbaceous plants that allows for the dispersal of the naïve juvenile mice, which results in less mouse-to-mouse contact. In contrast, during the fall, the dying vegetation reduces suitable habitats (i.e., tall grasses and crawling vegetation) in areas where herbaceous plants predominate, which results in more crowding of mouse populations. While aggression, including wounding, is believed to occur primarily during the mating season, the consequences of reduction in suitable habitats and influx of large numbers of young naïve mice during the fall likely lead to crowding and increased competition for space, aggression, wounding, and indirect transmission of HTNV to these young populations and subsequent increased potential for human transmission (Mackelprang et al. 2001, Olsson et al. 2002, Hinson et al. 2004). This not only results in an increased proportion of acute HTNV infections and increased viral shedding, but also increased viral shedding in a more confined area, which results in the spread of HTNV into the environment, increasing risks of HFRS infections. Subsequently, by December, when few HFRS cases are observed, established territories and very low reproductive rates (<1%) result in less movement and aggression, transmission of HTNV among mice, and a lower proportion of acute HTNV infections (T.A.K., unpublished data). Environmental conditions alone do not explain the very high proportion of HFRS cases (>80% of all cases) from September–December in the ROK, as relatively dry and dusty conditions occur from late September through much of June, when the wet monsoons greatly increase rainfall, usually from late June through August. While human activities (i.e., harvesting rice and other crops) increase during September–October, during the spring, agricultural practices (i.e., tilling and planting) occur without any observed increases in HFRS cases. In addition, cases of HFRS among military personnel who train throughout the year are observed primarily from September–November (T.A.K., unpublished data). The conclusion is that the increased number of human cases in the fall is due, at least in part, to the infusion of young naïve rodents in combination with the potential for increased movement to more favorable habitats (especially disturbed habitats) with reduced winter space-limiting groundcover.
While HTNV is shed throughout the life of A. agrarius, it has been proposed the shedding of HTNV in excreta is greatest in acute infections (Lee et al. 1981; J.W.S., unpublished data). To support this, HFRS cases among ROK populations are not correlated with the seasonal prevalence of HTNV Ab+ in A. agrarius (Klein et al. 2011). While HTNV Ab+ rates tend to be higher for disturbed or space-limited habitats than for larger areas, capture rates are generally lower (Sames et al. 2009, Kim et al. 2011). When these factors are taken into consideration for transmission potential, relatively undisturbed habitats had similar or higher densities of HTNV Ab+ mice based on capture rates (T.A.K., unpublished data). Thus, the prevalence of HTNV Ab+ mice and capture rates must be considered together, in combination with acute infections and levels of viral shedding, environmental conditions (dry and dusty), and human exposure to potentially contaminated dusts, to provide a more accurate estimate of the risk of human infections.
One of five M. minutus was HTNV Ab+ and this may be due to spillover of HTNV infections from A. agrarius through wounding, indirect transmission of virus in the environment, or the presence of a novel species-specific hantavirus. However, attempts to identify the hantavirus by RT-PCR from lung tissue was not productive. At other training sites, low numbers of M. fortis, M. musculus, and M. minutus were HTNV Ab+, while lung tissues were similarly negative by RT-PCR for hantaviruses (T.A.K., unpublished data).
HFRS is a disease of relative low frequency in humans and is dependent upon human behavioral factors that are difficult to incorporate in risk assessment models (Eisen et al. 2007). While monitoring rodent populations, HTNV Ab+ rates, and seasonal reproductive rates may provide some warning, the most effective means of reducing HFRS cases is limiting human contact with rodents and inhalation of dust with virus-laden rodent excreta (McCaughey and Hart 2000). Limited surveillance of large patches of tall dense grasses narrowly separated by barren ground where artillery are fired at FP-60 identified relatively high rodent populations (23.3% capture rates) and HTNV Ab+ rates (28.6%) subsequent to a U.S. soldier acquiring HFRS attributed to exposure at FP-60 during the fall of 2005 (O'Guinn et al. 2008, Song et al. 2009b). The proximity of these grassy islands to firing artillery may have resulted in increased dispersion of HTNV-infected mice as a result of territorial competition and rodent-to-rodent or indirect transmission of HTNV (Douglas et al. 2001, Sauvage et al. 2002, Hinson et al. 2004, Kallio et al. 2006, O'Guinn et al. 2008, Sames et al. 2009, Song et al. 2009b). This may have lead to aerosolization of virus-laden excreta during convoy, site establishment, and firing of artillery (back blast). Thus, without rodent-borne disease surveillance, avoidance of training activities during the period of highest recorded evidence of transmission of HTNV to humans is the most practical approach (McCaughey and Hart 2000, Muranyi et al. 2005).
HTNV surveillance at military training sites showed that HTNV is geographically diverse. This information is useful to identify the area of HTNV transmission, as military personnel often conduct training at several widely dispersed training sites during a 1–4-week interval (Song et al. 2009b). These data have provided for a more precise investigation of environmental (e.g., landscape and vegetation), biological (e.g., rodent population densities and seroprevalence of HTNV in A. agrarius), disease characteristics (e.g., incubation period), and soldier training activities (e.g., convoy and back-blast from firing artillery) leading to HTNV infections. The consolidation and modernization of multipurpose range complexes that modify dirt and gravel roads and other training components are designed to reduce dust and exposure to HTNV-contaminated dusts. However, training sites, for example, FP-10 and FP-60, still exist where habitat modification, for example, cutting vegetation to increase predation, is impractical. In these cases, reduced usage is warranted during periods of greatest risk based on rodent-borne disease surveillance.
Footnotes
Acknowledgments
We thank COL Brian Allgood (deceased) and COL Hee-Choon (Sam) Lee, 18th Medical Command, Seoul, Korea, for their support in conducting rodent-borne disease surveillance. We thank Ms. Suk Hee Yi, Force Health Protection, 65th Medical Brigade, Seoul, Korea, for conducting data analysis and GIS mapping, and MAJ Rex Bergren and LTC Min Ro, Chief, Laboratory Services, USAMEDDAC-Korea, Seoul Korea, for their support. We thank COL Ronald Smith, Commander, USAMEDDAC-Korea, LTC Mitchell Meyers, Chief, Force Health Protection and Preventive Medicine, 65th Medical Brigade, Seoul, Korea, and Dr. Joel Gaydos, Armed Forces Health Surveillance Center, Silver Spring, MD, for their support throughout the study.
Financial Support
Funding for portions of this work was provided by the Armed Forces Health Surveillance Center, Division of Global Emerging Infections Surveillance and Response System Operations, Silver Spring, MD, and the National Center for Medical Intelligence, Ft. Detrick, MD.
Disclosure Statement
The views expressed in this article are those of the authors and do not reflect the official policy or position of the Department of Defense, the Department of the Army, or the U.S. Government. No competing financial interests exist.