
Abstract
Bartonella are arthropod-borne, fastidious, Gram-negative, and aerobic bacilli distributed by fleas, lice, sand flies, and, possibly, ticks. The zoonotic Bartonella species, Bartonella henselae and Bartonella clarridgeiae, which are the causes of cat scratch disease and endocarditis in humans, have been reported from cats, cat fleas, and humans in Australia. However, to date, there has been no report of B. henselae or B. clarridgeiae in Australian wild animals and their ectoparasites. B. henselae and B. clarridgeiae were detected in fleas (Ctenocephalides felis) from red foxes (Vulpes vulpes), an introduced pest animal species in Australia, and only B. clarridgeiae was detected in blood from one red fox. Phylogenetic analysis of the ribosomal intergenic spacer region revealed that the B. henselae detected in the current study were related to B. henselae strain Houston-1, a major pathogenic strain in humans in Australia, and confirmed the genetic distinctness of B. clarridgeiae. The identification and characterization of Bartonella species in red foxes in the Southwest of Western Australia suggests that red foxes may act as reservoirs of infection for animals and humans in this region.
Introduction
The European red fox (Vulpes vulpes) occupies a wide variety of habitats across the continents of Europe, Asia, and North America. In the Southern Hemisphere, the red fox occurs only in Australia (Rolls 1984), where it was imported during the 19th century by English colonists for the purpose of hunting (Rolls 1984). However, this activity was not sufficient to keep red fox numbers in check, and they have since contributed to major population declines in a number of native Australian fauna species (Saunders and McLeod 2007). They are now considered a serious invasive pest species, and programs to eradicate the animals are conducted throughout Australia (Saunders and McLeod 2007). Previous research has discussed the distribution of some of their diseases and parasites (Glen and Dickman 2005).
Bartonella species have been reported in wild canids in North America and Europe, including coyotes (Canis latrans), gray foxes (Urocyon cinereoargenteus), and red foxes (V. vulpes) (Henn et al. 2009, Gabriel et al. 2009, Marquez et al. 2009); and the possibility that these canid species may act as Bartonella reservoirs has been discussed (Henn et al. 2009, Gabriel et al. 2009, Marquez et al. 2009). So far, only Bartonella rochalimae has been isolated or detected from red foxes (Henn et al. 2009). Bartonella spp. have also been detected in various flea species collected from the wild canids (Marquez et al. 2009), and fleas have been proposed to be major vectors of Bartonella spp. among wild canids (Henn et al. 2009, Gabriel et al. 2009).
Here, we report the first detection of zoonotic Bartonella species, including B. henselae and B. clarridgeiae, in both fleas (Ctenocephalides felis) and blood collected from European red foxes in southwest Western Australia.
Materials and Methods
Samples collection
Flea and blood samples were collected from red foxes in March 2010, in the areas surrounding the towns of Katanning (20–21st March) and Boyup Brook (27–28th March) in southwest Western Australia. An excess of 500 red foxes were shot by volunteers and farmers as a part of the “Red Card for the Red Fox” 2010 culling program coordinated by the Department of Agriculture and Food, Western Australia. Carcasses were brought to a central location for recording within 48 h, at which point, flea and blood samples were collected from 164 red foxes. Storage of red fox carcasses together during culling implied that fleas were able to move between carcasses, and as such it was not possible to identify individual fleas originating from specific red fox carcasses. A total of 151 fleas were randomly collected from 164 red foxes by using a flea comb or tweezers. Fleas were stored in 70% ethanol until they were processed for DNA extraction. Blood samples were collected from the peritoneal cavity of 14 red foxes into EDTA tubes and were stored at −20°C. Of these, ten were collected from red foxes in Katanning and four from Boyup Brook. Species of fleas were identified as C. felis by light microscopy using the standard key for Australian fleas (Dunnet and Nardon 1974). Between three and five fleas were pooled before DNA extraction, resulting in 34 pooled flea samples from the 151 fleas collected. Of the 117 fleas collected from Katanning, 25 flea pools were prepared and used for DNA extraction, whereas nine flea pools were prepared from 34 fleas collected from Boyup Brook (Table 1).
DNA extraction and PCR
DNA was extracted from pooled fleas and individual blood samples by using a DNeasy Blood and Tissue Kit (Qiagen) according to the manufacturer's instructions. Nested-PCR of the ribosomal intergenic spacer (ITS) region and the gltA gene was initially performed to detect Bartonella DNA in both flea pools and blood samples. External primers for the gltA gene and the ITS region were designed from the DNA sequences of B. henselae held in GenBank (gltA: L38987, ITS: L35101) (Table 2). The internal primers for nested-PCR of the ITS region and the gltA gene were as previously described, respectively (Norman et al. 1995, Jensen et al. 2000). An internal reverse primer targeting the ITS region was also designed for amplification and sequencing of a large fragment of the ITS region. DNA amplifications and sequencings of other loci including the 16S rRNA, ftsZ, and rpoB genes were performed to confirm the species status of Bartonella species detected in this study. All primers used in this study were described in a previous study (Kaewmongkol et al. 2011). PCR products from all genes were purified from agarose gel slices by using an UltraClean™ 15 DNA Purification Kit (MO BIO Laboratories, Inc.). Sequencing was performed by using an ABI Prism™ Terminator Cycle Sequencing kit (Applied Biosystems) on an Applied Biosystems 3730 DNA Analyzer, following the manufacturer's instructions. Nucleotide sequences generated for all 5 loci were analyzed by using Chromas lite version 2.0 (
ITS, intergenic spacer.
Results
Bartonella species were detected in 24 of the 34 DNA samples (70.5%) from pooled fleas using nested PCR of the ITS region and gltA gene. All 24 positive samples were from fleas collected from the area surrounding the town of Katanning (only one sample from fleas from this area was negative for Bartonella spp.). DNA sequencing of the 16S rRNA, gltA, ftsZ, and rpoB genes and the ITS region in all 24 Bartonella PCR positive samples revealed that 20 PCR positive samples were B. clarridgeiae, and 4 were B. henselae. A concatenated phylogenetic tree of all 5 loci was constructed to identify Bartonella species in this study (Fig. 1). B. clarridgeiae was also detected in 1 of the 10 blood samples collected from red foxes in Katanning (Table 1). The ITS region sequence of B. clarridgeiae amplified from the blood of this fox was identical to the corresponding B. clarridgeiae DNA sequences from fleas. Partial sequences for the five loci corresponding to these B. henselae and B. clarridgeiae detections were submitted to GenBank under the accession numbers HM990954, HM990959 (16S rRNA), HM990955, HM990960 (gltA), HM990956, HM990961 (rpoB), HM990957, HM990962 (16S-23S rRNA ITS), HM990958, and HM990963 (ftsZ). All of the sequences for a particular Bartonella species were identical across all samples amplified.
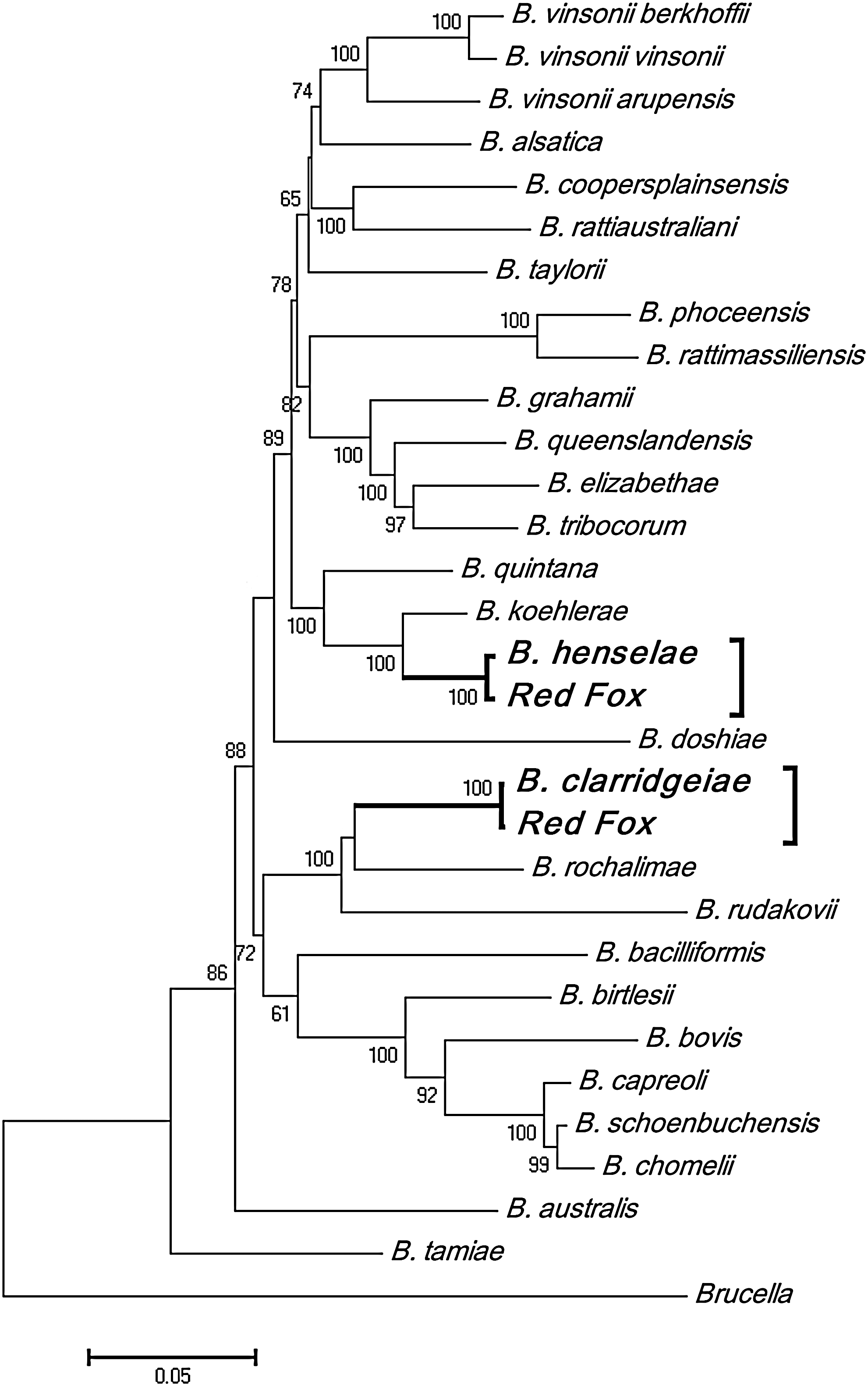
Neighbor-joining concatenated phylogenetic tree of 16S rRNA, gltA, ftsZ, rpoB, and the intergenic spacer (ITS) region of Bartonella henselae and Bartonella clarridgeiae detected in fleas from red foxes. Percentage bootstrap support (>60%) from 10,000 pseudoreplicates is indicated at the left of the supported node.
Phylogenetic analysis of B. henselae was conducted by using an 862-bp fragment of the ITS region. Distance analysis of the ITS region showed the close relationship between B. henselae detected in fleas from red foxes and B. henselae strain Houston-1 (0.3% genetic distance) (Fig. 2). An 893-bp fragment of the ITS region of B. clarridgeiae from the current study was also compared with other isolates of B. clarridgeiae by using distance analysis, and the resultant tree revealed that the B. clarridgeiae detected in red foxes was genetically distinct from previously published sequences of B. clarridgeiae (0.1%–2.8% genetic distance) (Fig. 3).

Neighbor-joining phylogenetic tree of the ITS region of B. henselae isolates. Percentage bootstrap support (>60%) from 10,000 pseudoreplicates is indicated at the left of the supported node. The tree is rooted by using Bartonella quintana as an outgroup (GenBank accession numbers AF368396).

Neighbor-joining phylogenetic tree of the ITS region of B. clarridgeiae isolates. Percentage bootstrap support (>60%) from 10,000 pseudoreplicates is indicated at the left of the supported node. The tree is rooted by using B. henselae as an outgroup (GenBank accession numbers L35101).
Discussion
This is the first report of B. henselae and B. clarridgeiae, two zoonotic species of Bartonella, from red foxes and their fleas in Australia; and this is the first time that B. clarridgeiae has been identified in a red fox. Until now, only B. rochalimae has been isolated or detected from red foxes from France (Henn et al. 2009). In the current study, concatenated phylogenetic analysis of all 5 loci confirmed the species status of B. clarridgeiae detected from fleas from red foxes, which exhibited 9.5% genetic distance from B. rochalimae (Fig. 1). Single-step PCRs for the 16S rRNA, ftsZ, and rpoB loci were unable to amplify B. clarridgeiae DNA in the blood of a red fox. However, B. clarridgeiae detected in the blood of a red fox was identical to the corresponding B. clarridgeiae sequences from flea extracts at the ITS region. Mixed sequences between B. henselae and B. clarridgeiae were not detected in this study.
B. clarridgeiae was detected in only one blood sample from a red fox. The true prevalence of Bartonella in the fox host may be higher than this result suggests; a pre-enrichment procedure followed by PCR detection has been shown to greatly improve the sensitivity of detecting Bartonella DNA in dog blood samples (Duncan et al. 2007). However, the red fox may not be the only natural reservoir for B. henselae and B. clarridgeiae in this region. Investigation of other wildlife reservoirs living in the same area, including native marsupials, feral cats, rabbits, and pet dogs, should be performed to further elucidate the ecology of the organisms.
The distance between the towns of Katanning and Boyup Brook in the southwest of Western Australia is approximately 120 kilometers. However, all positive samples were detected only in fleas collected from red foxes in the area of Katanning. It is not known why Bartonella species were not detected in flea samples from Boyup Brook, and further work needs to be performed to elucidate the ecology of Bartonella across the region. Multi-locus sequence typing has previously been conducted to differentiate B. henselae strains detected in cats and humans in Australia (Iredell et al. 2003). B. henselae sequence type 1 (ST 1), also known as strain Houston-1, has been identified as the principal strain causing human bartonellosis, and it is distributed widely in the cat population in Australia and North America (Iredell et al. 2003, Arvand et al. 2007). Distance analysis of the ITS region revealed that the B. henselae strains detected in the current study are closely related to B. henselae strain Houston-1 (ST 1) (0.3% genetic distance). The distribution of this strain in other mammalian hosts and their fleas should be defined, particularly in domestic cats in southwest Western Australia.
B. clarridgeiae detected in the current study seems to be distinct from other isolates using distance analysis of a large fragment of the ITS region. Although a prevalence study of B. clarridgeiae in cats and cat fleas from Eastern Australia has been previously reported (Barrs et al. 2010), there is little information on the ITS sequences of B. clarridgeiae in Australia in GenBank. Two separate ITS trees were produced due to the high level of variation at the ITS region, the phylogenetic analysis of B. henselae and B. clarridgeiae were resolved much better by conducting two separate analyses.
In conclusion, the identification and characterization of Bartonella species in red foxes in the SW of Western Australia suggests that foxes may act as reservoirs of infection for other animals, both wild and domesticated, and humans in this region.
Footnotes
Acknowledgments
The authors would like to thank Linda McInnes, Louise Pallant, Yazid Abdad, Michael Banazis, Rongchang Yang, Natasha Norrish, and Josephine Ng for their technical support.
Disclosure Statement
No competing financial interests exist.