
Abstract
Babesioses are hematic tick-borne diseases that induce malaria-like disorders in domestic, wild animals, and humans. Although indirect fluorescent antibody test (IFAT) and enzyme-linked immunosorbent assay (ELISA) commercial kits are available to test the presence of antibodies against most Babesia species, no kit exists to serologically diagnose the infections due to Babesia divergens, one of the most important zoonotic species. To fill this gap and to develop assays to detect animal and human infections, in vitro cultures (microaerophilous stationary phase system) of B. divergens were organized. Infected erythrocytes were adsorbed as corpuscular antigen (CA) on IFAT slides and ELISA microwells. The supernatant medium of the cultures (metabolic antigen, MA) was collected and employed in ELISA and western blot (WB) assays. B. divergens was also used to produce positive sera in Meriones unguiculatus and to infect a calf. Serological tests were set up with sera from experimentally/naturally infected animals, and possible cross-reactions were evaluated using heterologous sera from cattle positive to other piroplasms. Sera from clinically healthy people at risk of infection were also tested. As expected, assays based on the purified MAs from in vitro cultures proved more sensitive and specific than CA-IFAT and CA-ELISA. In fact, MA-ELISA provided satisfactory performances (even if 8.4%–15.7% cross-reactions were evidenced), and the WB developed proved totally sensitive and specific. WB indicated as immunodominant antigens two major protein bands at 33 and 37 kDa, which were also evidenced in 2.2% of the human sera tested, proving the parasite transmission to humans also in Italy.
Introduction
To fill this gap, we have developed IFAT, indirect ELISA, and western blot (WB) assays utilizing corpuscular antigens (CAs) and metabolic antigens (MAs) derived from B. divergens in vitro cultures to specifically detect animal and human infections.
Materials and Methods
In vivo and in vitro cultures
Research design, including animal experimental infections, was approved by the Italian Ministry of Health (protocol number: 16801-A
B. divergens strain Rouen, 1987 (kindly supplied by Prof. Carcy, Laboratoire de Biologie Cellulaire et Moléculaire, Montpellier, France), was intraperitoneally inoculated in Mongolian gerbils (Meriones unguiculatus) to have a source for the in vitro cultures (20 animals) and mainly to produce positive control sera (8 animals). Two gerbils were maintained as negative controls.
The isolate produced in gerbils was used for in vitro cultures using a microaerophilous stationary phase system on bovine and ovine erythrocytes (Bonoli et al. 2010). Briefly, RPMI-1640 medium (Sigma, St. Louis, MO) supplemented with 10% Gibco fetal calf serum (Invitrogen Ltd., Paisley, UK), 0.01% Gibco
The cultures, initially carried out on 24-well plates (200 μL of erythrocytes and 1 mL of medium each well) and subsequently on 6-well plates (800 μL erythrocytes+4 mL medium) and on 250-mL flasks (1200 μL erythrocytes+6 mL medium), were incubated at 37°C in a humidified 5% CO2 atmosphere. Every day they were fed by removing the medium overlying the erythrocyte layer and replacing it with fresh medium. Subcultures were carried out weekly by doubling, adding half the cultured erythrocytes to a new well or flask, and adding an equal amount of fresh erythrocytes. Once a week, parasitemia was checked by making Giemsa-stained erythrocyte smears.
A suspension of bovine infected erythrocytes (parasitemia 2.2%, a total of 3.7×109 infected cells) was used to infect a calf, whereas another calf was kept as negative control. Before the infection, both animals were tested for babesiosis by direct (blood smears and polymerase chain reaction [PCR]) and indirect (using commercially available IFAT and ELISA kits) methods. Their blood was collected every 2 days for 3 months after the infection and frozen at −20°C.
Ovine infected erythrocytes were used to obtain CAs and MAs for immunological assays described in this article.
Antigens preparation and coating
When parasitemia reached 10%, cultured ovine erythrocytes were centrifuged at 2800 rpm at 4°C, and the pellet was used as CA for IFAT slides and ELISA wells. Protein concentration was determined by Bradford assay as 5 ng/μL. Briefly, to prepare well slides for IFAT, the ovine erythrocytes were washed three times in phosphate-buffered saline (PBS) and packed by centrifugation; a volume of 10 μL was suspended in 4.17 mL of PBS. Each well was filled with 10 μL of the suspension (CA); the slides were dried, acetone-fixed, and stored at −20°C.
To prepare ELISA wells, polystyrene microplates were coated overnight at 4°C with CA (50 ng/well) diluted in 0.05 M sodium-carbonate–bicarbonate buffer (pH=9.6) and then stored at −80°C.
Culture supernatant was removed daily and then collected and stocked at −20°C. A volume of 50 mL was concentrated by polyethylene glycol (Sigma-Aldrich, St. Louis, MO) and dialyzed for 1 day at 4°C, against 1 L of 1 mM ethylenediaminetetraacetic acid (pH=8.0). The protein concentration of the solution (designated as MA), quantified by Bradford assay, was 17.3 μg/μL. This antigen was employed in ELISA and WB assays.
To prepare ELISA wells, MA diluted in sodium-carbonate–bicarbonate buffer (0.05 M, pH=9.6) was adsorbed (200 μL/well) for 1 h at 4°C; after three washings with PBS+0.05% (v/v) Tween-20 (pH=7.4), each well was blocked with 5% bovine serum albumin (BSA) in 1 M PBS and incubated at 37°C for 60 min; subsequently, the microplates were stored at −80°C.
As for WB, MA (50 μg/μL) was subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis for 2 h at 100 V and transferred onto nitrocellulose membrane (Osmonics) using semidry transblot (Bio-Rad, Life Sciences, Hercules, CA). The membrane was cut into strips, blocked for 1 h with 3% BSA in TBS at room temperature, then washed with TBS+0.05% Tween-20, and stored at+4°C.
Indirect fluorescence antibody test
IFAT was carried out on the slides carrying prefixed erythrocytes. Anti-B. divergens antibodies were detected by applying a standard procedure (Duh et al. 2007) and using rabbit anti-bovine IgG conjugated to fluorescein isothiocyanate (Sigma-Aldrich). Wells were counterstained with 0.02% Evans blue containing 30% glycerol and were submitted to fluorescence microscopic analysis at 40× magnification. Samples were classified as positive if fluorescence was observed at serum dilutions ≥1:40.
Indirect ELISA tests
Two indirect ELISA tests were developed based on different recognized antigens: parasitized erythrocytes (CA) and metabolic soluble antigens (MA).
The first assay was performed following the protocol described by Machado et al. (1997), which was slightly modified. In brief, sera diluted 1:20 in PBS-T containing 1% (v/v) of yeast extract were added to each well in duplicate, and plates were incubated at 37°C for 90 min. After three washings, monoclonal anti-bovine IgG1-HPR and IgG2-HPR (Sigma), diluted 1:5000 in PBS-T, were added at the same time to each well; plates were then incubated at 37°C for 60 min. TMB was applied as chromogen substrate. The enzymatic reaction was stopped by adding 0.5 M H2SO4. Absorbance was measured at 450 nm using an automatic ELISA plate reader (Bio-Rad). The cutoff value (0.350) was determined by the OD mean of 50 sera from healthy controls plus three times their standard deviation.
The second indirect ELISA test was performed using MA and applying the following protocol. Sera diluted 1:50 and secondary antibodies HPR (1:2000; anti-bovine, anti-human, or anti-rabbit for cattle, human, or gerbil sera, respectively; Sigma) were employed. Orthophenylene diamine (Sigma) in 0.05 M citrate buffer (pH=4.0) with 0.04% (v/v) H2O2 was added as substrate, and the enzymatic reaction was stopped with 0.5 M H2SO4. OD was measured at 492 nm (Bio-Rad, Life Sciences). The cutoff, determined with the aforementioned method, was established at OD=1.0.
Western blotting protocol
Sera, diluted 1:250 in the blocking solution, were added to individual strips and incubated at room temperature for 60 min. The strips were then washed five times and blots developed with antibodies (anti-bovine, anti-human, or anti-rabbit) conjugated with alkaline phosphatase (Sigma) diluted 1:2000 in the blocking solution at room temperature for 60 min. After five washings, strips were immersed in the substrate solution (BCIP/NBT-AP; Sigma), and the reaction was developed in the dark at room temperature for 15 min. A final washing step was performed once with distilled water.
Serum samples
In total, 491 sera were analyzed. IFAT, ELISA, and WB were set up with sera from experimentally infected gerbils (n=8) and calf (n=1), from bovines (n=4) naturally infected by B. divergens (diagnosed by PCR, using the primers AP1 and RLB-R2 that amplify a fragment of 800 bp from piroplasm 18S rRNA and following protocols described by Centeno-Lima et al. 2003 and Herwaldt et al. 2003), and from noninfected gerbils (n=2) and calf (n=1). Heterologous sera from cattle serologically positive to Babesia bovis (n=121) and Babesia bigemina (n=59) or molecularly positive to piroplasms including Theileria annae (n=15) and Theileria sergenti/buffeli/orientalis group (n=63) were employed to check any cross-reaction. Further, 50 sera from animals molecularly/serologically Babesia-free were included as negative controls. Each sample was screened for the presence of antibodies against CAs and MAs by IFAT and ELISA; positive or ambiguous reactions were checked by WB, as a sample of negative sera (n=20). Finally, 167 sera from people at risk of infection (90 breeders and 77 veterinarians) without clinical sign of babesiosis were also analyzed with MA tests.
Tests evaluation
The performance of each assay was evaluated as sensitivity, specificity, accuracy, positive predictive value, and negative predictive value. True and false positives, and true and false negatives, were calculated using sera from experimentally/naturally infected animals and negative controls.
Results
The results of the analyses carried out using serological assays developed with CAs and MAs are presented in Tables 1 and 2.
CA, corpuscular antigen; MA, metabolic antigen; IFAT, indirect fluorescent antibody test; ELISA, enzyme-linked immunosorbent assay; WB, western blot; pos.; ex, examined; exp, experimentally.
PCR, polymerase chain reaction.
Briefly, all the tests properly classified piroplasm-free animals. ELISA MA-based assay also showed high sensitivity (92.3%), whereas tests based on CA did not detect all experimental infections, with ELISA procedure proving more sensitive than IFAT (84.6% vs. 53.8%). Experimental WB correctly identified all positive sera from experimentally/naturally infected animals (sensitivity=100%), evidencing two major protein bands at 33+37 kDa. Accuracy of WB, MA-ELISA, CA-ELISA, and IFAT was 100%, 94%, 87%, and 62%, respectively, whereas NPV ranged from 33% to 100%.
Results concerning the screening of heterologous sera are reported in Table 2.
Tests developed with MA proved more specific than CA-based tests. In detail, IFAT misdiagnosed a lower number of B. divergens infections than CA-ELISA (which heavily cross-reacted mainly in the case of B. bigemina infections), but a higher number than MA-ELISA. As for WB, heterologous sera recognized protein patterns different from the 33+37 kDa displayed in experimentally/naturally infected animals. In fact, B. bovis- and B. bigemina-positive samples showed bands at 33 and 50 kDa, respectively, and the T. sergenti/buffeli/orientalis group at 30, 33, 39, or 50 kDa. A 37+50 kDa pattern was evidenced in animals positive to T. annae (Fig. 1).
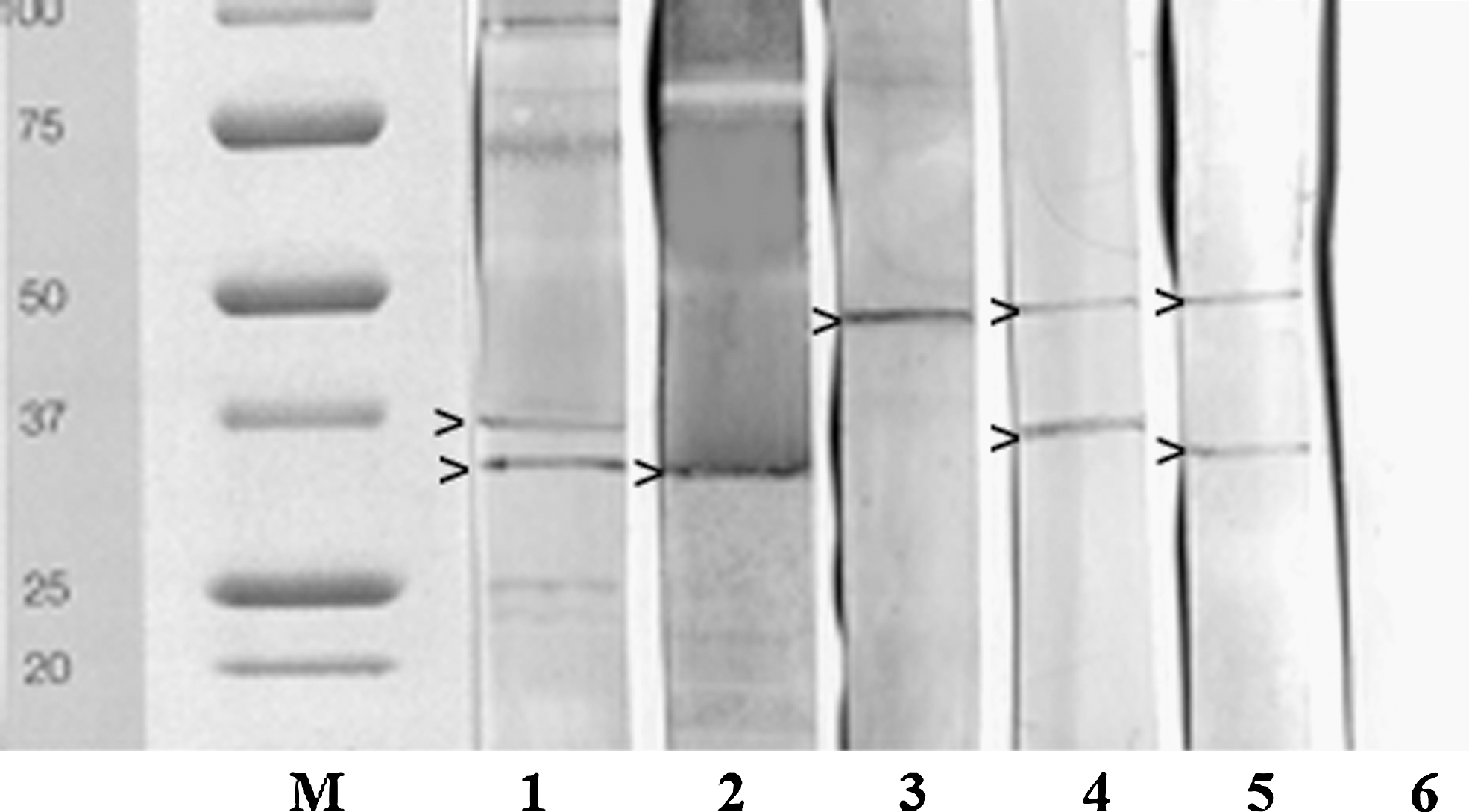
Protein patterns evidenced by metabolic antigen-western blot in sera from animals infected by Babesia divergens (33+37 kDa: lane 1), Babesia bovis (33 kDa: lane 2), Babesia bigemina (50 kDa: lane 3), Theileria annae (37+50 kDa: lane 4), and Theileria sergenti/buffeli/orientalis group (33+50 kDa: lane 5); lane 6=negative control from piroplasm-free subject; M=protein standard All blue (Bio-Rad).
Human sera were only submitted to MA-based tests. A total of 2/90 (2.2%) breeders showed anti-B. divergens antibodies, evidenced by ELISA and confirmed by WB; each anamnesis reported a tick bite event. No veterinarians proved seropositive to B. divergens.
Discussion
Unlike most of previous assays, the preliminary results of this study were obtained utilizing antigens from B. divergens in vitro cultures, a laborious method but one that allows the production of metabolic antigens, which are more specific than corpuscular ones. In fact, CA-IFAT and CA-ELISA tests showed sensitivity values of 53.8% and 84.6% and many cross-reactions in case of other Babesia infections. Therefore, even if incipient coinfections or previous infections cannot be excluded, the application of CA-based assays could produce false-positive results in areas where many Babesia species are sympatric. On the contrary, tests designed with MA gave satisfactory results. ELISA, in fact, detected the majority (92%) of experimental infections (but cross-reacted in 15.7%, 8.4%, and 14.1% of infections due to B. bovis, B. bigemina, and other piroplasms, respectively), and WB proved totally sensitive and specific (sera positive to B. divergens displayed a protein pattern different from heterologous sera). Therefore, although experimental MA-ELISA could be useful for a first-level screening, it must be followed by WB for the specific confirmation of positive sera. In addition, these results confirmed Bd37 as the major immunodominant antigen in B. divergens infections (Carcy et al. 1995), together with the 33-kDa protein, present also in B. bovis positive sera (Edelhofer et al. 2004). As Theileria species showed variable protein patterns (from 30 to 50 kDa), further positive samples should be tested to identify their immunodominant antigens.
The preliminary application of the tests to healthy people working in endemic babesiosis areas displayed anti-B. divergens antibodies in 2.2% of them. A slightly lower value was found in Germany in healthy blood donors (Hunfeld et al. 2002). This result suggests that the B. divergens transmission to humans also occurs in Italy, more frequently than diagnosed. Therefore, extensive serological trials are necessary to (i) quantify the prevalence of human infections in Italy and their current geographical distribution, (ii) identify the possible infection risk by each Babesia species, and (iii) evaluate the true sanitary relevance of these zoonoses. This means that studies necessary for the production of standardized commercially available diagnostic tools to detect animal and human infections due to B. divergens should be begun.
Further researches are currently in progress to challenge the specificity in cases of animals and people affected by other parasitic diseases, to develop a competitive inhibition-ELISA test, and to produce monoclonal antibodies to identify the immunodominant antigens of B. divergens-like strains that seem responsible for the majority of human infections in Europe.
Footnotes
Acknowledgments
The authors express gratitude to Prof. Carcy for providing the B. divergens strain, to Drs. C. Bonoli and S. Aureli for their skilful assistance in setting up the cultures, and to Dr. R. Cassini for his helpful suggestions. This research was supported by grants from the Italian Ministry of University and Scientific Research (Programmi di Ricerca di Rilevante Interesse Nazionale [PRIN] 2006).
Disclosure Statement
The authors disclose that there are no commercial associations and declare that no competing financial interests exist.