
Abstract
This review aims to summarize the current knowledge of the eco-epidemiology of Crimean-Congo hemorrhagic fever (CCHF) virus transmission reviewing the most recent scientific advances in the last few decades of epidemic and non-epidemic (“silent”) periods. We explicitly aim to highlight the dynamics of transmission that are still largely unknown. Recent knowledge gathered from research in Africa and Europe explains the very focal nature of the virus, and indicates that research on the ecology of the virus in the inter-epidemic periods of the disease has not yet been addressed. Hyalomma spp. ticks have been incriminated in the transmission of the virus under field conditions, but the role of other ticks found infected in nature remains to be tested under experimental conditions. Published evidence suggests that the increase in human cases reported in the Balkans, Turkey, and Russia is perhaps less due to the effect of changes in climate, but rather result from the impact of yet unexplored mechanisms of amplification that might be supported by wild animal hosts. Assessment of the available data suggests that epidemics in Eastern Europe are not the result of a spreading viral wave, but more likely are due to a combination of factors, such as habitat abandonment, landscape fragmentation, and proliferation of wildlife hosts that have exacerbated prevalence rates in tick vectors. There is an urgent need to empirically demonstrate these assumptions as well as the role of birds in introducing infected ticks, and also to evaluate the potential for survival of introduced ticks. Either a replacement of the pathogenic virus in the western Mediterranean or a lack of westward dissemination of infected tick populations may explain the absence of the virus in Western Europe.
Introduction
For other tick-borne diseases such as tick-borne encephalitis (TBE) and Lyme disease it has been speculated that changes in climate, host abundance, social habitats, economic fluctuations, environmental change, and to a lesser extent climate change, have increased the incidence of such diseases (Randolph 2001; Sumilo et al. 2007, 2009; Jaenson and Lindgren 2011). While the onset of the epidemic of Lyme disease in United States encouraged research on similar pathogens in Europe in the latter years of 20th century, the same interest has yet to be applied to other less proverbial tick-borne pathogens such as CCHF virus (CCHFV). This may in part be due to the relative lack of large CCHF outbreaks during the 1980s and 1990s in Europe and the former Soviet Union, coupled with the requirement for BSL-4 laboratories when handling live virus. The majority of contemporary research has been conducted in Turkey following the unprecedented epidemic in 2002. Further work has focused on endemic regions in South Africa, West Africa, Eastern Europe (Kosovo and Albania), and Central Asia (Tajikistan).
The complex enzootic cycles of CCHFV are made up of a combination of interactions resulting in a seemingly focal geographic distribution range which does not appear to currently match that of competent vector species. These interactions include: (1) several tick species implicated as vectors, (2) a wide array of (suspected) reservoir hosts for immature and adult stages of the ticks, (3) trends in climate, and (4) social changes, and consequently thereafter (5) alterations in landscape and vegetation. This review aims to summarize the most recent findings on CCHFV with a particular focus on the eco-epidemiology with a view to highlighting areas of importance where further research should be focused. Much knowledge has been gleaned in the mid-20th century, and will not be repeated here, including many basic aspects of the eco-epidemiology of the disease. The reader is specifically referred to Hoogstraal (1979). As well as pointing to recently acquired knowledge, we sought to stress missing basic factors in our understanding of the epidemics of CCHF, as well as some arguments that have already been rejected as drivers of re-emergence.
Tick species of importance
CCHFV is maintained in the field mainly by ticks of the genus Hyalomma. Some other tick species from the genera Dermacentor, Boophilus, Amblyomma, Rhipicephalus, and Haemaphysalis, have been found to harbor the virus in the field or have been artificially infected in the laboratory, but there is little evidence of their involvement in natural transmission or maintenance of foci (Watts et al. 1988). All reports linking the transmission of the virus by way of an infected vector have involved ticks of the genus Hyalomma (Watts et al. 1988). It would also appear that additionally Hyalomma ticks are also necessary for the maintenance of active foci of the virus in the field, even during periods of enzootic (or silent) activity.
The tick genus Hyalomma is widespread in different ecological areas of the Palearctic and Afrotropical regions. Some species, like Hyalomma scupense (one- or two-host biology) and H. anatolicum (two- or three-host biology) prefer to feed on the same large ungulates (mostly cattle) during all developmental stages, and adopt a nidicolous life cycle. For Hyalomma marginatum and H. rufipes the life cycle is a two-host one, with immature ticks feeding upon birds or small/medium-sized mammals, and after molting, adults feed upon a larger host (wild and domestic ungulates) (Apanaskevich 2004). The principal species implicated in transmitting CCHFV in Eurasia are H. marginatum, H. turanicum, H. anatolicum\, and H. scupense (including the former H. detritum, now considered a synonym of H. scupense; Guglielmone et al. 2009). The virus has been reported to survive throughout the life cycle of the tick and passes trans-stadially and trans-ovarially. The long survival of the virus in ticks is important from an epidemiological point of view. However, there is still a dearth of knowledge regarding host exposure rates and host immune responses, particularly in populations of short-lived birds, insectivores, and lagomorphs. Such animals have a high population turnover shown to be important in other tick-borne pathogens (e.g., TBE virus), where such hosts develop antibodies to exposure in the nest during their first few days of life.
The epidemiological potential, relating climate, ticks, and reservoirs of the active foci, is a very important part of the enzootic ecology of CCHFV. Hoogstraal (1979) reported up to 69 tick species as infected under laboratory or natural conditions with CCHFV. However, the involvement of these species in the epidemiology of the virus under natural conditions seems less clear. First, there is a lack of clinical cases in regions where some of the tick species involved in transmission are widespread. This is particularly evident in Kosovo, where H. marginatum ticks are widespread with little obvious difference in available hosts and habitat in regions where CCHFV is reported compared to regions where it is not. Similar findings have been reported in Turkey, and significantly in other countries such as Spain, where CCHF cases have not yet been reported in spite of the presence of potential tick vector species.
Previous studies in Tajikistan (Pirov et al. 2005) have revealed an interesting pathway of endophilic foci. In the peridomestic environment of the residential farms in southern Tajikistan, both H. anatolicum and H. scupense are abundant, and it is not uncommon to find 100 feeding adults on the same host. Since both species may complete their life cycles within the protection of the walls and crevices of barns, ticks may be very abundant without any other host but livestock. After the examination of 7414 Hyalomma spp. ticks, collected during the years 2002 to 2004, by a nested-PCR using combined pools of ticks, 80 pools of H. anatolicum were found to be positive for CCHFV (out of 710), and none of H. scupense (out of 33 pools). The highest number of positive ticks was observed when feeding specimens were examined, possibly indicating a viremic state of the hosts on which the ticks were collected. Similar observations were made in Turkey, where egg batches and larvae obtained from female H. marginatum ticks fed on cattle, and were found positive for CCHFV RNA (Vatansever et al. 2008). Such findings highlight the need to reassess the role of large wild and domestic ungulates, commonly considered insignificant in virus amplification, but known to be significant in supporting tick populations by feeding the adults. For several species restricted to cattle such as H. scupense, H. anatolicum, and Rhipicephalus annulatus, which have been found to be positive in several CCHFV-endemic countries, it must be presumed that these “refractive” hosts are indeed responsible for infecting feeding ticks, or possibly that trans-ovarial rates of transmission, and/or co-feeding occurs at a higher rate than previously thought. Given a lack of recent studies using more sensitive and user-friendly techniques such as RT-PCR and ELISA, the role that previously reported refractive hosts play in the natural CCHFV life cycle is unclear.
Lessons from CCHFV studies in Africa
CCHF was diagnosed for the first time in South Africa by isolation of the virus from the blood of a 13-year-old boy who died after attending a nature study camp (Swanepoel et al. 1983). The furthest south the virus had previously been found in Africa was Tanzania. The virus was isolated previously in countries from Senegal in the west, to Kenya in the east (Hoogstraal 1979). The field investigations, which followed recognition of the disease, included antibody sera collected from humans and livestock, and a survey of the prevalence of the virus in questing and feeding ticks. Collection of questing ticks in the surroundings of the study camp reported adults of H. rufipes and H. truncatum. Up to 21 isolations of the virus were successful from the adult ticks, and the prevalence of the virus was found not to be significantly different in the two species (Swanepoel et al. 1983). Adult Hyalomma ticks collected on Taurotragus oryx (common eland), and one adult Rhipicephalus evertsi (out of 12) collected on sheep were also positive, but not Margaropus winthemi collected on the same animals. The few immature ticks collected were negative. Sera from Lepus spp. (hares, 4/10), Bos taurus, and Ovis aries (cattle and sheep, 183/440) reacted positively to a reverse passive hemagglutination inhibition test. However, the viremia of CCHFV in artificially infected cattle was not sufficiently high to infect adults of H. rufipes (Rechav et al. 1987, citing a personal communication by R. Swanepoel). Sera from hares collected in sites at Cape Province, Natal, Transvaal, and Free State in 1977–1979 were also positive (28/161) (Swanepoel et al. 1983). These findings suggested that the virus is widespread in South Africa, and perhaps the lack of human cases as seen in Eurasia is due to the lower pathogenicity of the African strains of the virus compared with the European and Asian ones.
Further studies were carried out in west Africa, mainly in Senegal and Mauritania (Wilson 1990; Chapman et al. 1991; Zeller et al., 1994a, 1994b, 1997). These studies highlighted the clear correlation between antibodies to the disease in livestock and humans, and the distribution of ticks of the genus Hyalomma (Wilson et al. 1990). Rainfall in Senegal varies from 200 mm in the Sahelian zone in the north, to more than 1400 mm in the sub-Guinean zone in the south, and this is reflected in the changing composition of the tick species across the country. Bioclimatic zones differed in the intensity with which CCHFV was transmitted. Evidence of infection in sheep was greatest in the northern, arid, sparsely-vegetated zone of Senegal, and decreased consistently toward the southern, moister, forested zone. The tick vectors that maintain CCHFV transmission in Senegal are unknown (Wilson et al. 1990), although their results indicated that Hyalomma species are important in the maintenance of local or regional foci of the disease. Further studies (Sylla et al. 2008) focused also on the clear clinal transition of climate variables along a north-south gradient in Senegal as a marker for the dominant tick species, and therefore the serological prevalence of CCHFV in humans.
Lessons from the re-emergence of the disease in Eurasia
The ongoing epidemic of CCHF in Turkey began with some isolated cases in Tokat province (Gozalan et al., 2004). The human health authorities soon realized that more clinical cases were being reported from neighboring sites, and then later over a large portion of the country (Yilmaz et al. 2009), largely coinciding with the expected distribution of the tick H. marginatum (Estrada-Peña et al. 2007). This was demonstrated in an early article about the dynamics of the infection in that country (Estrada-Peña et al. 2007). The very focal nature of CCHF in Turkey exhibited a strong correlation between the highest incidence in human cases with the presence of particular land use categories, significantly small and highly mosaic agricultural localities. This was consistent with the most common transmission mechanism reported, the bite of an infected tick, and with the increased densities of ticks in fragmented sites, which in turn provide an environment for human exposure.
Similarly to other tick-borne diseases, climate trends have been commonly linked to outbreaks of clinical cases. Ticks are highly sensitive to small changes in climate, as long-term, sustained, and small differences in key climate variables may drive a serious change. However, without a framework linking the response of the tick to subtle climate variations, it is only possible to speculate in the case of CCHFV. An assessment of the effects of climate on the presence of human clinical cases of the disease in Turkey was carried out (Estrada-Peña et al. 2009). The analysis included monthly values of several climate variables and concluded that climate was no different in sites with active foci of the disease compared with sites where H. marginatum is common but human cases are not reported. They concluded that it is not possible to predict, based solely on climate grounds, where new cases would appear in the near future. Climate is not the sole factor driving the virus amplification, even if climate in autumn/winter may have a strong regulating role on the survival of tick populations (Hoogstraal 1979). It is noteworthy that Hyalomma-endemic areas with the mildest autumns and winters in the Mediterranean basin have historically been, and are currently free of the disease, so the impact of a warmer climate appears not to be the only factor affecting the occurrence of the disease. Efforts to link disease occurrence solely to an array of climate conditions (Randolph and Rogers 2007) may be unrealistic. Thus claims that a warmer climate influences the prevalence of the disease (Purnak et al. 2003; Vorou et al. 2009) are unsupported when climate series are examined and compared. Bold statements such as “the probability of hospitable climates not only extended to Spain, Italy, and northern African countries across the Mediterranean sea, but even in large parts of North America, smaller parts of South America, and Australia” (Maltezou and Pappa 2010) are unreliable without further knowledge of the system. Even if climate may be consistent with sites where the disease has been reported, adequate vectors for CCHFV are known to be absent in the New World and Australia. The role of the tick vectors needs therefore to be carefully evaluated under both laboratory and field conditions, and the lack of concordance between both groups of data clarified.
There is, however, evidence that a fragmented landscape, with many small patches existing within a matrix of unsuitable tick habitat, may lead to isolated populations of both ticks and hosts, producing an amplification cycle whereby ticks feed on infected hosts (Estrada-Peña et al. 2011). For CCHFV eco-epidemiology, the degree of habitat patchiness contributes to the increased contact rate among reservoir hosts, humans, and ticks. It also leads to the occurrence of isolated amplification foci, with consequently higher tick exposure to humans (Estrada-Peña et al. 2009). Isolated fragments surrounded by a matrix of agricultural land commonly have the poorest diversity of mammals. Species composition changes within fragmented and smaller isolated patches, and each species differs in its ability to cross the matrix between fragments (Pires et al. 2002; Lira et al. 2007; Forero-Medina and Vieira 2009). Although Hyalomma ticks can move horizontally, they require a vertebrate host to move over significant distances. Because these host populations are relatively isolated, there are few local movements of hosts, and therefore new naive animals carrying uninfected ticks are not diluting the prevalence rates in the isolated patch; however, the immune response of such isolated populations against the ticks and the pathogen may seriously decrease the population of infected ticks. These aspects have never been studied for CCHFV.
The epidemic in Turkey is unlikely to be caused by a spreading wave of infected ticks by dispersing hosts with an “imported” virus strain (or by a new, more infective genetic variant of the virus) spreading from central Anatolia to other regions of Turkey, as was hypothesized by Karti and associates (2004). Research on the epidemiology of the CCHF infection in Turkey has focused on the region of Anatolia, north-central Turkey, due to the high incidence. The first clinical case of CCHF in the Thrace region (in northwestern Turkey) was noted in 2007 (Midilli et al. 2009). Results from a serosurvey of 193 human samples from three localities showed that IgM and IgG rates are compatible with an epidemiological situation in which the virus has been present for a long time, and is not the result of a recent invasive event from the main epidemic center in Anatolia (Gargili et al. 2010). The detection of CCHFV IgM positivity and an elevated proportion of IgG seropositivity indicated that the genetic variants Europe 1 (the clade in which viral strains from Anatolia are included) and Europe 2 are very active in the region, and that high IgG rates in humans indicate active immune protection (Gargili et al. 2010). These figures support the idea that the infection was present in Turkey for a long time before the epidemic started. Conditions have existed for continued cycles of amplification, and only recently the epidemic threshold was reached, and this is not the final step in a chain of recent climate trends. Such an epidemic threshold, whatever the factors behind it, has been observed simultaneously in distant regions of Turkey in only 2–4 years, and is not the result of a moving wave of infected ticks feeding on hosts. While landscape patchiness, and thus isolated amplification systems, seems to be one of the causes, the factors driving the enzootic amplification cycle in Turkey remain unknown.
In both Turkey and Kosovo, changes in animal husbandry might also be important with respect to the hare population and its associated tick infestations. In the last 20–30 years the sheep and goat populations have decreased by 50% in Turkey (observations by Z.V.), and 65% (sheep), 75% (goat) in Kosovo (unpublished data; Faculty of Agriculture Kosovo 2007), with this decrease more pronounced in places where CCHF has been reported. Hares avoid pastures grazed by sheep and prefer cattle pastures (Smith et al 2004), therefore a decrease in the sheep population may have contributed to an increase in hares. Land use practices are also important; the agriculture in CCHF areas is based on crop rotation land fallowing, and in a given village during half the year land is used for wheat production, and for the remaining months is used for either chickpea/vetch production or is left fallow. The following year the land use is rotated. This farming strategy creates habitat heterogeneity, which is ideal for hares. Such changes in arable strategy also impact the dominant tick species. For example Rhipicephalus spp. dominate when sheep populations are high, with an increase in H. marginatum associated with a decrease in sheep and increases in hare and cattle.
Two of the authors (L.J. and J.M.) studied the recent rise of clinical cases of CCHF in Kosovo. Several driving forces behind the rise in incidence were identified, and these broadly include: (1) a decline in agriculture, (2) an expansion of grazed grassland, (3) scrub encroachment close to villages, (4) an imbalance in local predator-prey cycles, and (5) a shift from local ecto-parasite-resistant cattle breeds to naïve European breeds (Jameson et al., in prep, a). This combination of driving forces is thought to not only have led to an increase in Hyalomma ticks, but also elevated the contact between such ticks and humans, as well as an overall increase in the proportion of Hyalomma ticks that are infected.
The R0 and why it is difficult to obtain for CCHF
One of the most useful notions in infectious disease epidemiology is the case reproduction number, R0, defined as the average number of secondary cases produced by a typical primary case in an otherwise fully susceptible population. R0 is of central importance because its value determines whether an epidemic is possible; only when it takes a value greater than one, so that each primary cases generates on average more than one secondary case, can the chain reaction that constitutes an epidemic continue. When R0 is less than 1 there may still be chains of disease transmission, and occasionally these may even be quite long, but these will be self-limiting and have no chance of leading to the self-sustaining chain reaction of a full-blown epidemic. The well-known equation for insect-borne infections is:
N/H is the ratio of vectors of host, a is the daily biting rate, the coefficient β expresses the transmission rates from vertebrates to insects and vice-versa, p is the vector daily survival probability, h the host daily mortality, and r the daily rate of loss of infectivity in the host.
However, R0 is a concept conceived for insect-borne diseases that has been transferred to tick-borne diseases. Conclusions drawn on the transmission rates extrapolated from insects to ticks are not always true, and the modes of transmission of tick-borne pathogens have shown to be particularly complex. The particular life history of ticks has an impact on the transmission rates, or, inversely, some factors widely acknowledged to be of extreme importance in the insect-pathogen system rarely have an impact in a tick-pathogen system. The reader is referred to the review by Randolph (1998) for the biological reasoning behind the very different dynamics between tick-borne and insect-borne diseases. Randolph (1998) proposed the following equation for tick-borne parasites:
Where total R0 is the sum of the partial R0s for each tick stage and each species of host, f is the probability of a tick feeding on an individual of a particular host species, and F is the vector reproduction rate. Randolph (1998) mentioned that the off-host interstadial period, comprising both post-engorgement development and questing for the next host, is functionally equivalent to a very long extrinsic incubation period (n), as the tick is not infective until it is ready to feed again. Furthermore, transmission potential by ticks is very sensitive to climatically-induced variations in tick development rates (Randolph 1998). Other than the long developmental periods, different rates of horizontal and vertical transmission, and a large array of hosts available to feed different stages of the vector, tick-borne pathogens have a very important feature: the enhanced transmission by a co-feeding route. For some pathogens with short infectious periods in hosts, the infection of ticks by systemic transmission seems much less efficient than for pathogens with longer or more chronic infections. The co-feeding transmission between infected and uninfected ticks that feed in close proximity on non-viremic hosts, has been demonstrated to occur in the CCHFV life cycle (Gordon et al., 1993), and has been proposed as they key pathway that can sustain infections by some viruses in ticks, like TBE virus (Labuda et al., 1993). Studies by Nonaka and associates (2010) using a comprehensive modeling framework showed that under conditions of tick inter-cohort overlap, the co-feeding route may be the most important method of transmission of a pathogen among the tick vectors. That route may have different relative importance when different vertical transmission rates are simulated.
Hoogstraal (1979) highlighted the impact that changes in wildlife hunting practices can have on the amplification of CCHFV and/or ticks. This has recently been identified as a potential driver in Kosovo, where hunting was heavy in the early 1990s, and was then banned in 1993 with little illegal hunting (ARD-USAID 2003). As a result the numbers of hares, wolves, and wild boar increased dramatically in the early 2000s. Hares are known to be important hosts for immature Hyalomma ticks, and are proficient amplifiers of the virus (Camicas et al. 1990). Discussions with local hunters in Kosovo (by L.J. and J.M.) suggested that there had been a noticeable increase in tick numbers on local hare populations, as well as a local increase in hare numbers. The latter was attributed to a decline in the hare's main predator, the red fox. A similar situation has also been recognized by Z.V. in Central Anatolia (Turkey). Further field surveys that investigate the infestation rates of different wild animals (mammals and birds), and the infection rates within the animals and their ticks, would facilitate a better understanding of transmission cycles. It would also help us understand the impact that hunting might have in keeping important amplifier species at threshold levels within endemic areas, so as to impact transmission cycles and R0 of the virus.
The relationship between Hyalomma ticks and CCHFV in the recent epidemic foci of Eastern Europe likely depend on complex interactions of multiple ecological, environmental, and societal variables. Factors that contribute to an increase in the abundance of the hosts for immature ticks would likely have an impact on R0, since such hosts would feed a higher number of ticks. It is explicitly assumed herein that density-dependent mortality of ticks is not increased in this scenario, or at least the feeding ticks are below a threshold over which mechanisms of host immunity would regulate the tick population. Hyalomma marginatum is a two-host tick, therefore larvae and nymph feed on the same animal. Results from a modeling framework concluded that the unengorged adult is the most common overwintering stage (Estrada-Peña et al. 2011), suggesting that the population is regulated by the mortality of molting nymphs in winter. An inter-cohort overlap (and hence a possible amplification by co-feeding) in tick populations could be expected if engorged nymphs could survive the winter in high numbers and complete their molt after the spring temperature rise. Owing to the lack of knowledge of the reservoir role of the hosts in the field, no attempts have been made to evaluate the potential importance of different parts of the viral infection in CCHFV. Recent proposals to estimate R0 (Foppa 2005; Hartemink et al. 2008) might help in the computation of that index for CCHFV. In addition to the role of climate on tick biology, it is crucial that other factors, such as the impact of habitat fragmentation and patchiness, are considered when deriving R0.
What is the role of migratory birds?
Migratory birds have been repeatedly implicated in dispersing immature Hyalomma. Each spring many thousands of ticks are introduced in Europe through migratory flights from Africa (Hoogstraal et al. 1961, 1963). The level to which these birds may be exposed to African Hyalomma populations and subsequently import them into Europe depends on the habitats they frequent, their ground feeding behavior, and the timing of their departure. Owing to the earlier timing of bird arrivals in Western Europe compared to the period for optimal nymphal molt in Africa, the likelihood of such importation is reduced (Knudsen et al. 2007; Estrada-Peña et al. 2011). The northern limit of their potential survival will depend on how suitable the abiotic conditions are at the arrival site to facilitate molting of engorged immatures, as well as the availability of suitable hosts. Hasle and colleagues (2010) reported the presence of only seven immature H. rufipes on 713 migratory birds collected in the southern coasts of Norway; however, this does not guarantee their survival under local climate conditions. Further investigation is required to determine whether or not such exotic tick species can survive and establish if they are fortunate enough to drop off in a suitable habitat and with available hosts. Hyalomma rufipes (from sub-Saharan Africa and the Nile River) cannot survive in the environments of the Mediterranean basin nor the northern European latitudes. Studies demonstrated that the natural distribution of H. rufipes is Sub-Saharan Africa and adjacent regions of Africa and Arabia along the Red Sea (Apanaskevich and Horak 2008). The records of H. rufipes from Europe (Macedonia, Malta, and Turkey), and North Africa (except Egypt), are an apparent consequence of the dissemination of the immature stages by migratory birds from Africa, but thus far do not represent permanent populations (Apanaskevich and Horak 2008). However, studies have demonstrated that the northern distribution limit of H. marginatum, which is a Palearctic species, may be moving further north owing to the trend in autumn and winter temperatures (Estrada-Peña and Venzal 2007). The issue is thus the evaluation of the probabilities that a bird with infected and feeding Hyalomma immatures may arrive at a given site in Europe, and the precise timing for optimal tick molt and survival, given that the trans-ovarial transmission route for CCHFV may infect around 1% of larvae.
Other than the eco-epidemiological role played by birds in the dispersion of ticks, previous studies have indicated that birds might be reservoirs of CCHFV, but most of the Russian studies suggest that birds are refractory to experimental inoculation with the virus, and are unable to re-isolate the virus, and thus have no serological evidence of infection (Hoogstraal 1979). However, viremia and antibodies were demonstrated in guinea fowl following CCHFV inoculation (Sheperd et al. 1987), indicating that the previous methods outlined by Russian workers were inadequate for antibody detection in birds. Another factor that may contribute to the negative serological results is the transient nature of the antibody production, as reported in guinea fowl following experimental CCHFV inoculation (Sheperd et al. 1987). Studies by Zeller and associates (1994b) of the role of ground-feeding birds as reservoirs for CCHFV in Senegal were conclusive about the presence of antibodies and immature Hyalomma ticks on several species of ground-feeding birds. CCHFV antigens have been found in the brains of rooks (Corvus frugilegus) in Rostov (Russia), where these birds are common hosts for immature H. marginatum (Moskvitina et al. 2004; Vodyanitskaya et al. 2008). The last factor to include in the equation is the lack of apparent circulation of the virus in northern Africa, which is the southern limit of the distribution range of H. marginatum, and is where birds begin the last step in their flights into Europe with immature H. marginatum ticks. Before we can quantify the role of migratory birds, further studies on the occurrence of the virus (and consequent infection) in northern Africa are necessary, as well as an assessment of the fate of the fed ticks introduced in Europe. Current U.K. studies by Jameson and colleagues (in prep, b) have highlighted that infestation rates of H. marginatum on migratory birds are higher than previously suspected.
Studies of the phylogenetic affinities of the viral isolates showed that the transmission and circulation of the virus is largely compartmentalized within Eurasia and Africa (Burt and Swanepoel 2005). Moreover, H. marginatum is a Palearctic tick, distributed north of the Sahara desert, and in the western coast of Africa, in countries including Senegal and Mauritania (Camicas et al. 1990; Apanaskevich and Horak 2008). Other species of Hyalomma, other than the nidicolous H. scupense and H. anatolicum, have not been reported in southwestern Europe or northern Africa (Estrada-Peña et al. 2004). It is important to note that ground-feeding birds, common hosts for H. marginatum, commonly fly over short distances along the Atlantic African coast to avoid the dangers of flying over the large Saharan desert and the lack of food and water resources (Sokolov et al., 1999). This may prevent the introduction of engorged immatures above an outbreak threshold because they would drop on the ground in the frequent stops the birds make for resting. The scenario of birds dispersing the infected ticks and expanding the CCHF foci could perhaps be adopted from rook and quail (Coturnix coturnix) movements, at least in Turkey. Instead of south-to-north introduction scenarios (African origin), east-west or north-south intercontinental bird movements may be more important (European origin to neighboring areas).
Why there may be a lack of human clinical cases in western Europe
One of the most striking questions in the geographical distribution of CCHFV is the lack of clinical cases in the western Mediterranean, west of the main distribution area of the pathogen, which ends at the western Balkans. This is of special interest because the main tick vector (H. marginatum) is common and abundant in places like Spain and southern Italy, as well as Mediterranean Africa (e.g., Morocco and Tunisia), where neither the disease nor the virus have been reported. Preliminary unpublished findings by the author (A.E.P.) from Spain have concluded that H. marginatum feeds on humans and in some localities may reach high densities. It is difficult to comprehend how the relative densities of each of key host species might influence the transmission cycle, and it may be that low densities of key hosts may explain the apparent absence of virus.
Figure 1 shows the compared distribution of both tick reports and CCHF human cases in the western Palearctic, as published by EFSA (2010), with an evidently greater range of the main tick vector compared to the disease, with human cases restricted to a subset of the vector range. Reports pointed towards a surprisingly high level of diversity of the viral strains (Hewson et al. 2004). However, the large amount of sequence diversity found within the virus argues against a recent origin of the virus (Carroll et al. 2010), and supports the hypothesis that CCHFV is an old virus that has been circulating for centuries. The long evolutionary history of CCHFV and Hyalomma ticks has provided ample opportunity for expansion to their current geographic ranges throughout Africa, Europe, the Middle East, and Asia, and it seems reasonable to conclude that the virus has already reached its full geographic extent where suitable conditions currently exist for its survival. Reports of new human cases at the beginning of the 21st century could be referred to as a re-emergence. There have been no reports of human cases in countries that have not previously detected human seroprevalence (Ahmed et al. 2009). Statements like “given the presence of suitable vectors, hosts, and climate parameters in south Europe Mediterranean countries, it appears possible for CCHF to emerge in these areas within the near future” (Maltezou and Pappa 2010) are unsupported, as in a system at equilibrium the virus should have already spread to those areas where the vector and the hosts have historically been present (Hewitt 2004). However, we cannot be sure that the virus is not existing in hidden enzootic cycles (e.g., in Hungary and Portugal where there is seroprevalence), and that the right combination of events may lead to the emergence of transmission to humans. Active surveillance should be undertaken to detect the virus in humans, animals, and ticks, where the tick's main vector exists.
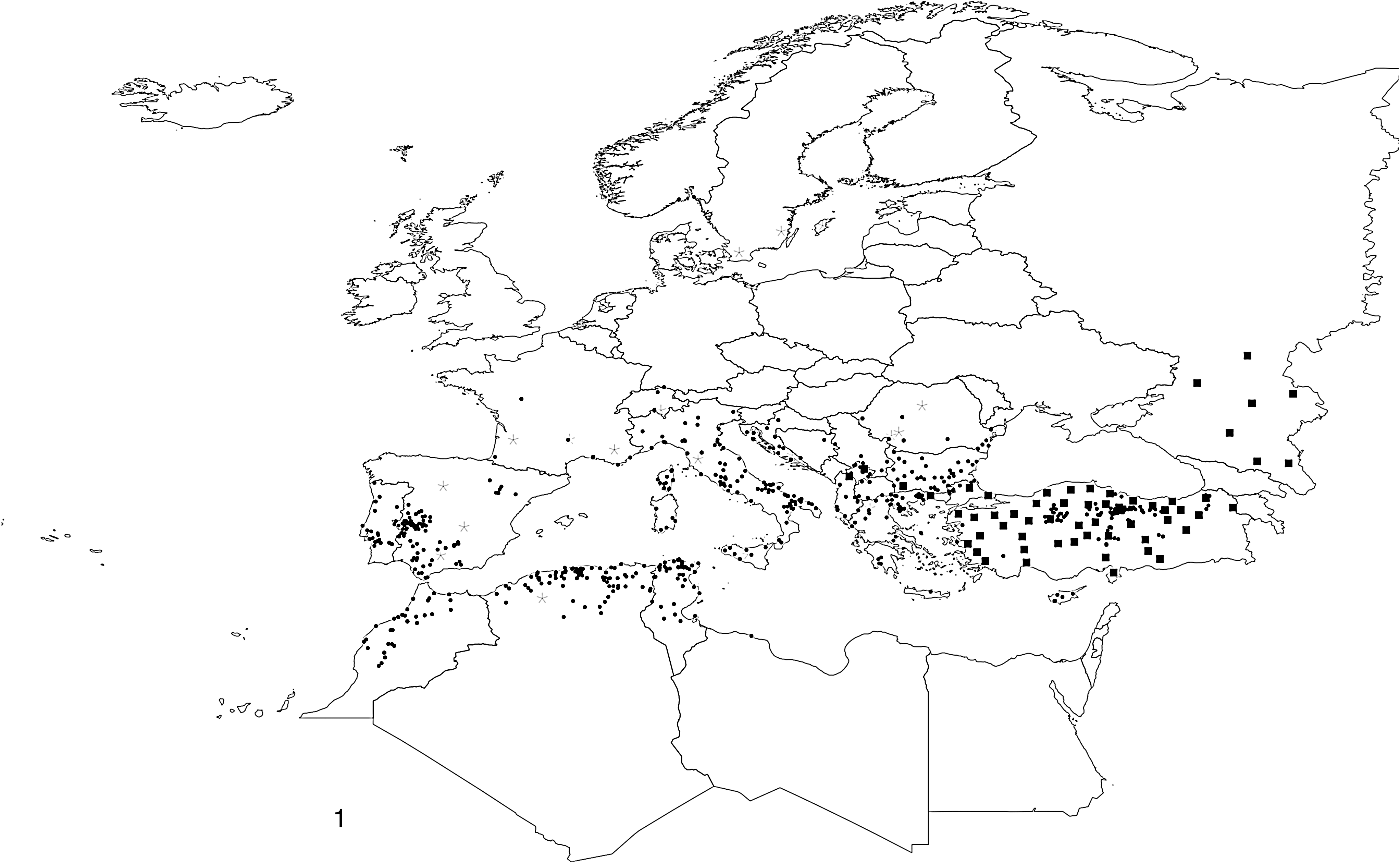
Map showing the reported distribution of Hyalomma marginatum and clinical cases of Crimean-Congo hemorrhagic fever (CCHF) in the western Palearctic, as published in the EFSA journal (2010). Dots represent records of the tick with reliable coordinate pairs (geo-located), and stars indicate records of the tick with reports of the administrative region (not unambiguously geo-located). Squares represent the centroid of the administrative region where cases of CCHF have been reported (EFSA 2010).
It is interesting to speculate that the lack of known cases from the western Mediterranean could be ascribed to one or several of the following hypotheses: (1) the strain of the virus present in the area has a low pathogenicity, (2) the virus is not reported in the western Mediterranean because the regional strains of the tick are not able to transmit the virus, and (3) the virus has not yet been reported in the western Mediterranean because there is a limited degree of spread of the infected eastern populations of H. marginatum towards western areas. We explicitly exclude the lack of adequate reservoir-amplifying hosts, and the lack of adequate climate niches, based on studies of the role of livestock as amplifying hosts (Hoogstraal 1979) everywhere they have been scrutinized. We will review these hypotheses below.
Regarding the first hypothesis, the existence of isolates of the Europe 2 genogroup of CCHFV in Greece (AP92) and western Turkey (KMAG) must first be considered (Gargili et al. 2010). These strains have a low pathogenicity and have clear phylogenetic separation from other CCHFV strains (Iashina et al. 2002; Midilli et al. 2009; Mild et al. 2009; Ozdarendeli et al. 2010). As previously mentioned, studies in the western part of Turkey bordering the region where the AP92 strain was first detected demonstrated that the joint distribution of both the Europe 1 (pathogenic) and the Europe 2 (non-pathogenic, but see Ozkaya et al. 2010) clades of the virus suggest immune protection. The lack of adequate serosurveys in western Europe, other than the one by Filipe and associates (1985), together with the absence of severe clinical features of infections by AP92-like strains, could be responsible for the lack of reported infections in such areas. It is interesting to note that the lower-pathogenic viruses found in Greece are phylogenetically close to known sequences from western Africa, in accord with comments about the low pathogenicity seen for the west African isolates (Swanepoel et al. 1983).
The second hypothesis relates to the presence of specific molecular receptors in the tick that facilitate transmission of the virus, an aspect of the ecology of the virus that has never been addressed. While pioneering studies have reported on the infection of tick organs by the virus (Hoogstraal 1979), there are no further insights into the way in which the virus attaches to tick cells and their receptors, and how viral particles are delivered to the rest of the tick's body cavity. Considering the variety of species in the genus Hyalomma, for which transmission has been proven in studies under controlled conditions, it seems improbable that the populations of H. marginatum present in western Europe lack an adequate array of biochemical signaling agents needed for successful viral replication and transmission.
The lack of interaction between eastern and western populations of H. margimatum is an exciting explanation for the apparent absence of clinical cases in the western Mediterranean. This hypothesis is based on the probable absence of mixing of infected eastern and uninfected western populations of the tick. The apparent border between these populations presumably is west of the Balkan endemic region, where limited animal movements prevent western spread of the tick. It remains unclear why the Europe 2 clade is found in parts of the Balkans and western Turkey, but have been reported only once in the eastern portion of the country (Ozkaya et al. 2010), which has been subjected to intensive sampling. If unrestricted movements of ticks exist between central Turkey and the Balkans, then the Europe 2 clade strains should be found in central Turkey. This could be further explained by trade regulations operating in western Turkey, near the Balkans, where livestock movements are restricted. Furthermore, wild animals, like Sus scrofa and Lepus spp., have a distribution bottleneck in western Turkey (Larson et al. 2005; Kasapidis et al. 2005). The hares collected in Turkey have been demonstrated to be genetically isolated from the species in Europe west of the Greece-Turkey border (Kasapidis et al. 2005). In addition to the isthmus of Istanbul, that acts as a natural bottleneck to animal movements (and hence ticks), a hypothetical northern route for the spread of Hyalomma ticks may also be constrained by colder climates that would prevent tick colonization (Estrada-Peña et al. 2011). The lack of trade movements of livestock, coupled with the apparent lack of movements of wild hosts for ticks, contribute to a general lack of connectivity between eastern and western populations of the vector and virus. The Europe 2 (non-pathogenic) clade would thus remain in Europe west of the Balkans, and the Europe 1 clade east of the Balkans, with a small area of overlap by diffusion near Greece and Thrace. We postulate that southwestern Europe may be free of clinical cases of CCHF because of both the probable presence of strains of the Europe 2 clade, and the lack of flow between the Near East tick and host populations and the European ones. Nonetheless further investigation of the drivers of emergence and studies looking at the seroprevalence in at-risk countries is advisable.
Conclusions
The eco-epidemiology of any tick-borne pathogen transmission cycle is complex. This is due to the changeable and adaptive life cycle of the involved tick vectors, which is further complicated by the direct impacts of climate on arthropod populations and the dynamics of the reservoir hosts. This reasoning is also of applicability to CCHFV, for which several basic features remain to be elucidated. One of the primary goals of this review was to discuss a series of facts implicitly accepted after the publication of previous reports, and to examine the dynamics of the virus in ticks and vectors that remain largely unknown. These accepted claims are unsupported by empirical proof and thus should no longer be considered in further reports until more evidence is available. The role of birds in the establishment of ticks in new areas (and not only their spread), the rates of antibody production in animals (as a diagnostic tool for infection rates), and the impact of single factors like climate or host abundance on the epidemiological chain must still be disaggregated and studied before their impact can be fully elucidated. The passionate and sometimes idealized claims about the spread of the viral infection because of single factors driving a small portion of the epidemiological chain of factors must be examined only after we have acquired an adequate body of knowledge about transmission dynamics. It is of extreme importance to understand if autochthonous infections exist in the Mediterranean basin and the phylogenetic relationships of the viral strains. An empirical background examination of the reasons for the absence of disease should be developed and evaluated. An epidemiological predictive map of risk would be the logical final step in this chain.
Footnotes
Acknowledgments
Parts of this article were the result of the inspiration of the people engaged in the understanding of the ecology of CCHF in Eastern Europe. A.E.P., Z.V., L.J., and J.M. wish to acknowledge the funding by ArboZoonet, an International Network for Capacity Building for the Control of Emerging Viral Vector Borne Zoonotic Diseases, project number 211757, under the European Union FP7 framework. A.E.P. also wants to acknowledge the discussions with Aysen Gargili (University of Marmara, Turkey).
Author Disclosure Statement
No competing financial interests exist.