
Abstract
West Nile disease (WND) has become a major public and veterinary health concern since the appearance of West Nile virus (WNV) in New York in 1999. The following panzootic spread in the U.S. and the recent WNV outbreaks in Europe and the Mediterranean Basin have increased interest in WND. Despite considerable investigation of WNV infection in birds, the effects of WNV on avian populations are still largely unknown. In Europe, raptors have been found to be particularly susceptible to WNV infection, but studies in birds of prey are lacking. To our knowledge, the present study is the first to report an experimental infection with WNV in Gyr-Saker hybrid falcons. We show that 10-week-old captive-reared Gyr-Saker (Falco rusticolus×Falco cherrug) hybrid falcons are susceptible to WNV infection. Neither morbidity nor mortality was observed after subcutaneous WNV inoculation with mixed extracts of non-infected mosquito salivary glands. Both the macroscopic and microscopic lesions observed were similar to those previously reported in naturally and experimentally infected North American raptors. The results obtained in the present study demonstrate that although Gyr-Saker hybrid falcons do not seem to be a good reservoir for WNV transmission via mosquito, they can become infected with WNV, develop viremia and antibodies, and are able to shed the virus.
Introduction
WNV belongs to the Japanese encephalitis serogroup within the Flaviviridae family (Weissenbock et al. 2010). It is the most geographically widely distributed arbovirus and two lineages of WNV are widely recognized (Lanciotti et al. 1999). Lineage I includes WNV strains from Africa, the Middle East, Europe, India, Australia (formerly Kunjin virus), and the Americas (Charrel et al. 2003). Until recently, all lineage II isolates were from sub-Saharan Africa and Madagascar, but between 2004 and 2008, it has been isolated from birds of prey in Hungary (Bakonyi et al. 2006; Erdélyi et al. 2007; Wodak et al. 2011) and was also detected in a Eurasian magpie in Greece in 2010 (Valiakos et al. 2011).
Since 1999, a dramatic spread of WNV occurred in the U.S.; over 25,000 human cases of WND have been reported (Murray et al. 2010). In the 1999 outbreak, fatal WND characterized by meningoencephalitis and myocarditis was observed in infected wild and captive birds (Steele et al. 2000; Nemeth et al. 2006a). However, not all birds died from infection with the New York 1999 (NY-99) strain, and these birds developed humoral immunity (Komar et al. 2001; Nemeth et al. 2008). Since then, hundreds of bird species have tested WNV positive, with American crows having the highest rate (Bernard et al. 2001). Native North American raptor species (including Falconiformes and Strigiformes) also showed high WNV susceptibility, and have been proposed as being useful for WNV surveillance (Joyner et al. 2006; Nemeth et al. 2007a,2007b,2009). As judged by magnitude and duration of viremia from experimental infections with the NY-99 strain, raptor species were considered as competent reservoirs for WNV (Komar et al. 2003). Moreover, raptors may become infected after consumption of WNV-infected hosts (Garmendia et al. 2000; Komar et al. 2003).
The WNV epidemiology in the Old World differs from that of the U.S., as it causes sporadic WND cases in humans and horses in Europe and the Mediterranean Basin (Húbalek and Halouzka, 1999). Avian mortality associated with WNV is rare in the Old World (Dauphin et al. 2004; Zeller and Schuffenecker 2004 ; Jourdain et al. 2007), although significant numbers of storks and domestic geese died during the epizootics in Israel (Malkinson et al. 2002). A recent experimental work demonstrated clinical susceptibility of European red-legged partridges to WNV (Sotelo et al. 2011). Moreover, fatalities due to WNV in raptor species, such as sparrow hawks and goshawks in Hungary (Bakonyi et al. 2006; Erdelyi et al, 2007), and in imperial eagles in Spain (Jiménez-Clavero et al. 2008), have been reported.
WND has become a major public and animal health concern following the appearance of WNV in New York in 1999, the unprecedented subsequent panzootic, and the recent WNV outbreaks in Europe and the Mediterranean basin (Calistri et al. 2010). Raptors are recognized as important reservoirs of this viral infection. Most wild birds of prey are protected in Europe, so captive-reared Gyr-Saker hybrid falcons were used as an experimental animal model for WNV infection. Therefore the aim of the current study was to evaluate the effect of the WNV NY-99 strain in Gyr-Saker hybrid falcons to assess whether they are susceptible to WNV infection.
Materials and Methods
Animals
Six 10-week-old male captive-reared Ɛgyr/¼ saker (Falco rusticolus×Falco cherrug) hybrid falcons were obtained from a breeder. From 3 weeks until 10 weeks of age the falcons were imprinted by a person from the research group in order to minimize further stress. One week prior to infection, antiparasitic treatment (toltrazunil) and hematologic tests were performed. The hematologic tests were normal. The birds were tested for the presence of antibodies against WNV using a competitive ELISA (cELISA). The falcons were tagged with aluminum leg bands and kept in isolator cages located in negatively pressurized isolation rooms within Biosafety Level 3 (BSL3) facilities at the Centre de Recerca en Sanitat Animal (CReSA), where they were acclimatized for 5 days. Whole prey was provided twice a day. Animal care and all procedures were performed in accordance with the ethics committee regulations of the Universitat Autònoma de Barcelona.
Virus
WNV strain NY-99-flamingo 382-99 was propagated in Vero cells after four previous passages in BHK-21 cells (Cordoba et al. 2007), and its complete sequence was obtained (Martín-Acebes and Saiz 2011). The virus for the inoculum was titrated by 50% tissue culture infective dose (TCID50) in Vero cells.
Experimental infection design
In an attempt to mimic natural infection, the falcons were inoculated with an inoculum made up of Culex pipiens salivary gland extracts mixed with WNV. Four falcons were subcutaneously inoculated with 0.1 mL of 4 log10 TCID50 of WNV strain NY-99-flamingo 382-99 mixed with the homogenate of 50 salivary glands dissected from 3- to 5-day-old Culex pipiens adult females. Taking into account that each mosquito has 2 salivary glands, the use of 50 salivary glands is expected to be equivalent to the saliva present in 25 adult mosquitoes, the number of mosquitoes that could bite a raptor in an area with a high density of mosquitoes. Two falcons were subcutaneously inoculated with the homogenate of 50 salivary glands resuspended in 0.1 mL of PBS, as a negative control group.
Clinical signs were daily recorded up to 14 days post-inoculation (dpi). Birds from the infected group were euthanized with intravenous sodium pentobarbital (100 mg/kg) and sequentially necropsied at 5, 7, 9, and 14 dpi. The two birds from the control group were euthanized and necropsied at 14 dpi.
Sample collection
Blood samples and oral, cloacal, and ocular swabs were collected daily until 9 dpi and at 14 dpi. Blood was obtained from peripheral veins (nail) or from the heart after anesthesia with ketamine/xylacaine (10 mg/kg Imalgene 1000 plus 1 mg/kg Xilagesic 2%). To obtain sera, the blood samples were left to clot prior to separation by centrifugation 6000 g for 5 min, and the separated sera were collected. One hundred microliters of serum was mixed with 0.4 mL of DMEM with 10% glycerol. Oral, cloacal, and ocular swabs were taken and soaked in 0.5 mL of DMEM with 10% glycerol. Samples of cerebrum, medulla oblongata, spinal cord, liver, spleen, kidney, and feather pulp were collected during necropsy, and 0.1 g of each tissue was introduced in 1 mL of DMEM plus 10% glycerol. All samples were maintained at −80°C until further testing for virus detection. Serum obtained from each animal after euthanasia was stored at −80°C without DMEM with 10% glycerol until further testing for seroneutralitzation. Samples of the above-mentioned tissues, as well as bursa of Fabricius, bone, heart, lung, skeletal muscle, small and large intestines, pancreas, trachea, esophagus, thyroid, adrenal gland, gonads, proventriculus, ventriculus, adrenal glands, eye, and sciatic nerve were collected and fixed in 10% buffered formalin for histology studies.
Viral quantitation by quantitative real time RT-PCR (RT-qPCR)
WNV loads in oral, cloacal, and ocular swabs, sera, and homogenized tissue samples were assessed by a quantitative fast TaqMan one-step RT-qPCR using Fast7500 equipment (Applied Biosystems, Carlsbad, CA). Viral RNA was extracted with a QIAamp viral RNA mini kit (Qiagen, Valencia, CA). Amplification of a 3′NC region fragment was performed with primers and probes as previously described (Jiménez-Clavero et al. 2006), at concentrations of 900 nM for each primer and 200 nM for each probe, using fast AgPath-ID one-step RT-PCR reagents (Applied Biosystems). The amplification conditions were: reverse transcription at 48°C for 10 min, an initial denaturation reaction at 95°C for 10 min, and 45 PCR cycles of 97°C for 3 sec and 61°C for 30 sec.
Standard curves and quantification were achieved by amplification of a 75-bp fragment of the 3′NC region using the aforementioned NY-99 strain as a template following the amplification conditions described above. The obtained 3′NC region fragment amplicon was cloned into pGEMT vector (Promega Corp., Madison, WI), and transformed in Escherichia coli-competent cells (Invitrogen Corp., Carlsbad, CA). The recombinant plasmid was purified using the QIAprep Spin kit (Qiagen), and quantified using the fluorometer Qubit (Invitrogen), following the manufacturer instructions to obtain the concentration of the plasmid. The copy number of recombinant plasmids was calculated as previously described (Fronhoffs et al. 2002). Serial 10-fold dilutions of plasmid of known concentration were made, and the standard curves were generated using 2.45×101 to 2.45×106 copies of recombinant plasmid. The limit of detection (LoD) for the NY-99 strain was 24.53 plasmid copies per reaction, which corresponded to 0.014 TCID50 per reaction. The genome equivalent copies (GEC) of plasmid from the collected samples were determined based on these standard curves and taking into account their volumes.
Virus titration
Ten-fold dilutions in PBS of the samples were spiked onto PBS-washed Vero cell monolayers grown in 96-well microtiter plates. Eight wells were infected for each 10-fold dilution, and 20 μL of inoculum was spiked to each well. The plates were incubated at 37°C and 5% CO2 for 1 h, followed by the addition of 150 μL of DMEM supplemented with 2% fetal bovine serum, 2 mM L-glutamine, 100 UI/mL penicillin, and 100 μg/mL streptomycin (Sigma-Aldrich, St. Louis, MO). The plates were then incubated at 37°C and 5% CO2 for 7 days. Viral titration was calculated using the Reed and Muench method.
Antibody detection
Sera from all falcons were tested for the presence of WNV antibodies by a WNV commercial cELISA (ID Screen® West Nile Competition; IdVet, Montpellier, France). Sera from euthanized falcons at 5, 7, 9, and 14 dpi were tested for WNV neutralizing antibodies by means of a micro-virus-neutralization test (VNT) as previously described (Figuerola et al. 2008).
Histopathology and immunohistochemistry
Standard paraffin-embedding histological procedures and hematoxylin and eosin staining were used to prepare the formalin-fixed tissues for microscopic examination. For immunohistochemistry, 3-μm paraffin sections were deparaffinized and rehydrated, and incubated with 3% hydrogen peroxide in methanol for 30 min at room temperature for endogenous peroxidase inhibition. The sections were incubated with protease 0.1% (protease type XIV bacterial from Streptomyces griseus; Sigma-Aldrich) for 8 min at 37°C for antigen retrieval, followed by incubation with Tris-buffered saline with 2% bovine serum albumin (Sigma-Aldrich) for 1 h at room temperature to block unspecific staining. The sections were incubated with the primary antibody, a rabbit anti-WNV polyclonal antibody (1:500; BioReliance, Rockville, MD) at 4°C overnight, followed by incubation with secondary antibody biotinylated polyclonal goat anti-rabbit immunoglobulin (1:200; DakoCytomation, Carpenteria, CA) for 1 h at room temperature, and avidin-biotin complex (ABC Peroxidase Staining Kit; Thermo Scientific, Madrid, Spain) for 1 h at room temperature. The reaction was developed by incubation with 3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich) for 5 min, followed by counter-staining with Mayer's hematoxylin. The positive control was formalin-fixed paraffin-embedded tissue from a WNV-infected specific pathogen-free chicken embryo. For a negative control, duplicate sections were incubated with rabbit negative control serum (Negative Rabbit Serum; BioReliance).
Results
Clinical signs and WNV viremia profiles in falcons
No clinical signs were observed during the period of experimental infection (14 days). All falcons survived the infection and developed viremia. WNV-RNA was detected by RT-qPCR in serum of all infected falcons as early as 2 dpi. Viral load could be detected from 2–9 dpi, peaking at 4–6 dpi (Fig. 1). The highest WNV-RNA load in serum of individual infected falcons ranged from 7.19–7.59 log10 GEC/mL (Fig. 1). Infectious virus was detected from 2–7 dpi (Fig. 1). WNV serum titers around 4 log10 TCID50/mL persisted on average for 4 days from 3–6 dpi, after which the infective virus decreased drastically.
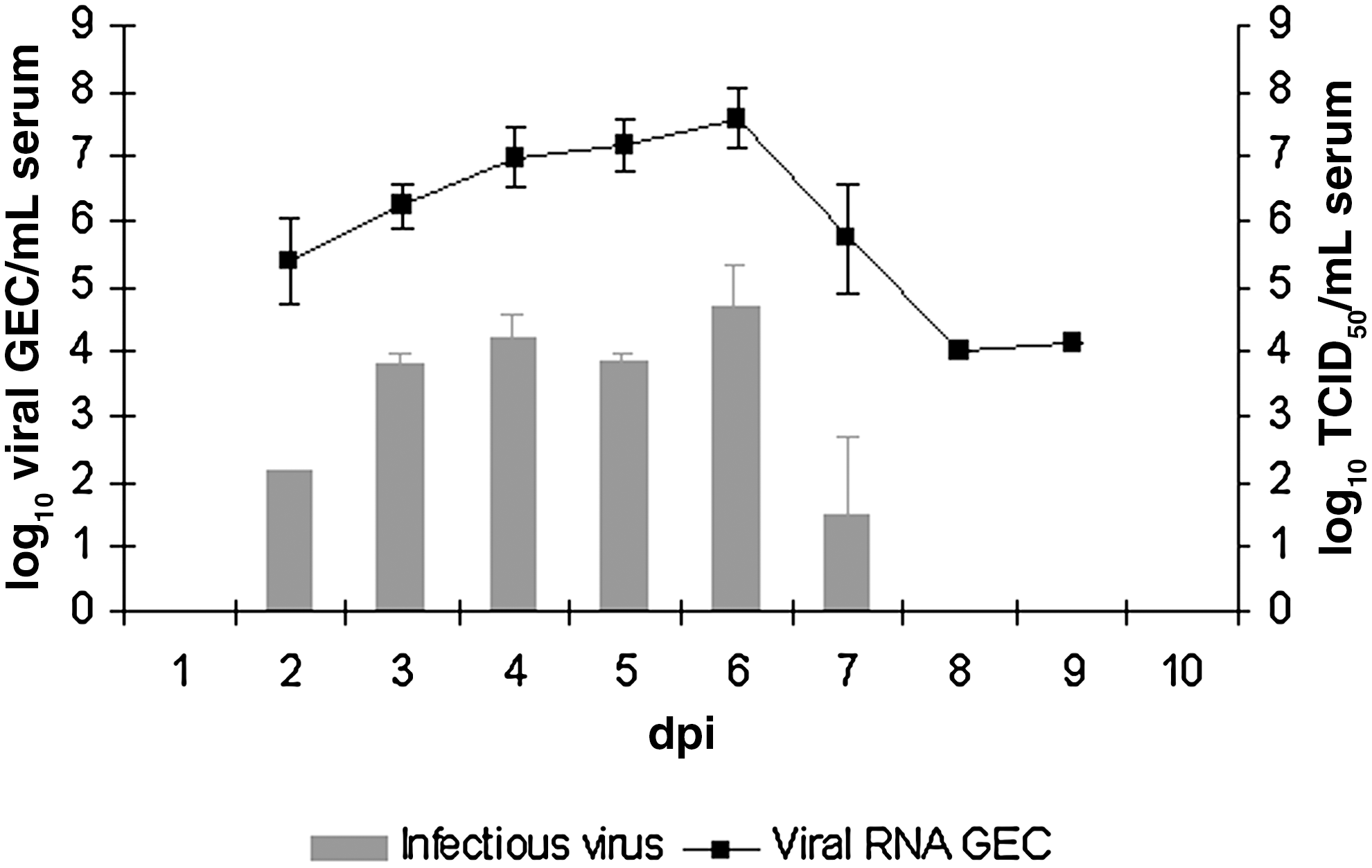
Viral load expressed as log10 viral genome equivalent copies (GEC) and log10 50% tissue culture infective dose (TCID50) per 1 mL of serum at different days post-inoculation (dpi). The standard deviation is indicated by error bars.
WNV shedding patterns in WNV-infected falcons
WNV shedding patterns from oral, cloacal, and ocular swabs detected by RT-qPCR throughout the experiment are summarized in Table 1. Shedding of WNV was first detected from all three swabs at 3 dpi. Viral shedding extended beyond the period of viremia, with titers ranging from 3.42–6.66 log10 GEC up to 14 dpi in oral and cloacal swabs, and to 11 dpi in ocular swabs, although at very low levels. Only WNV from the oral and cloacal swabs obtained at 5 and 7 dpi was able to grow in Vero cells. All the swabs collected from the control group were negative for the presence of WNV.
Gyr-Saker falcons were inoculated with 0.1 mL of 4 log10 50% tissue culture infectious doses of WNV NY-99 strain mixed with the homogenate of 50 salivary glands dissected from Culex pipiens.
Ct (cycle threshold) value.
log10 viral genome equivalent copies/swab.
Infectious dose in log10 TCID50/swab.
n.d., no data.
WNV detection in tissues by RT-qPCR and virus isolation
All the tissues collected from the infected falcons at the necropsies were WNV-positive by RT-qPCR (Table 2). The falcon euthanized at 7 dpi had the highest viral load in spinal cord, liver, spleen, kidney, and feather pulp. In contrast, the falcon examined at 14 dpi showed higher viral RNA levels in the cerebrum and medulla oblongata than the other falcons and than any other tissue from that falcon.
Gyr-Saker falcons were inoculated with 0.1 mL of 4 log10 50% tissue culture infectious doses (TCID50) of WNV NY-99 strain mixed with the homogenate of 50 salivary glands dissected from Culex pipiens.
Infectious dose in log10 TCID50/g.
GEC, genome equivalent copies.
From all WNV RT-qPCR-positive tissue samples, WNV could be isolated from cerebrum, medulla oblongata, spinal cord, and feather pulp at 7 dpi, from cerebrum and spleen at 9 dpi, and from cerebrum at 14 dpi, and all of them had more than 3.19 log10 TCID50/g (Table 2).
Pathology and viral detection by immunohistochemistry
At necropsy the only macroscopic lesion observed was splenomegaly, which was present in all WNV-infected falcons. Microscopic lesions were more evident as the WNV infection progressed and are summarized in Table 3. The most striking lesions were observed in the central nervous system and heart of the birds necropsied at 9 and 14 dpi, and consisted of non-suppurative encephalitis and myocarditis. Neither macroscopic nor microscopic lesions were observed in the control falcons.
Gyr-Saker falcons were inoculated with 0.1 mL of 4 log10 50% tissue culture infectious doses of WNV NY-99 strain mixed with the homogenate of 50 salivary glands dissected from Culex pipiens in the lateral neck.
−, negative; +, mild; ++, moderate; +++, severe.
Scarce WNV antigen was observed by immunohistochemistry in all the infected falcons; however, the antigen distribution varied as the infection progressed. WNV antigen was observed in dendritic cells/macrophages of the spleen (at 5, 7, and 9 dpi); Kupffer cells, epithelial cells of renal tubules, corticotrophic cells of the adrenal gland, and lamina propria of the small intestine (at 7 and 9 dpi); lamina propria of the large intestine and cecal tonsil (at 7 dpi); and epithelial cells of the renal medullary tubule and tunica media of coronary arteries (at 14 dpi).
Serologic responses against WNV
Total serum anti-WNV envelope protein (pr-E) antibody levels were measured with a cELISA. All birds were seronegative at the beginning of the experiment. The results represented in Figure 2 show that seroconversion occurred at 7 dpi, and then the antibody levels increased until the end of the experiment at 14 dpi. Birds from the control group remained negative throughout the experimental infection. Moreover, the infected falcons developed neutralizing antibodies at 7 dpi, with reciprocal of the highest dilution VNT values of 20, which increased to 480 by 9 dpi, and decreased to 40 by 14 dpi (Fig. 2).
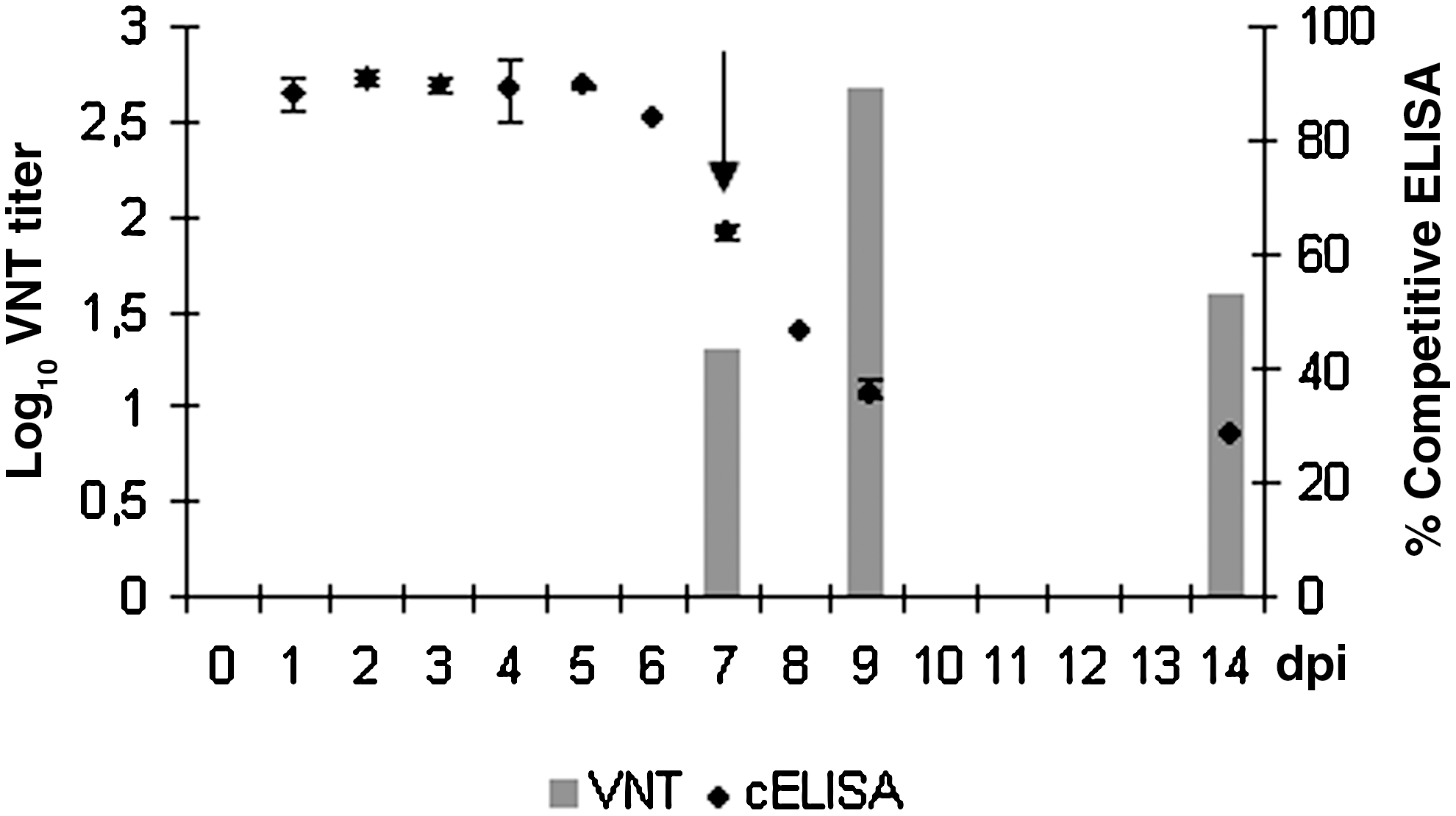
Detection of antibodies against WNV NY-99 strain by competitive ELISA (cELISA) and virus-neutralization test VNT. The arrow indicates the first day of antibody detection, at 7 dpi.
Discussion
The principal aim of the present experimental infection study was to assess the susceptibility of reared Gyr-Saker hybrid falcons to WNV to use them as an experimental animal model for WNV infection since other falcon species are protected in Europe. To our knowledge, this is the first experimental infection study with WNV in Gyr-Saker hybrid falcons. Among raptors, falcons are probably one of the most popular and extended group of species used in falconry, an art which is widespread, mainly in Arab countries.
The results of the present work demonstrate that Gyr-Saker hybrid falcons are susceptible to WNV infection and develop viremia and antibodies, although no clinical illness was observed. However, the limited number of Gyr-Saker hybrid falcons used in the present experimental infection does not allow us to infer whether WNV is pathogenic or not in a relevant percentage of falcons. Previous studies have reported morbidity after natural WNV infection in birds of prey, such as golden eagles (Aquila chrysaetos), red-tailed hawks (Buteo jamaicensis), great horned owls (Bubo virginianus) (Nemeth et al. 2006a, Hofle et al. 2008), Iberian imperial eagle (Aquila adalberti), Bonelli's eagle (Hieraaetus fasciatus) (Hofle et al. 2008; Jimenez-Clavero et al. 2008), goshawks (Accipiter gentilis), sparrow hawks (Accipiter nisus) (Bakonyi et al. 2006), and gyrfalcons (Falco rusticolus) (Wodak et al. 2011). However, the results of our experimental infection are in agreement with previous experimental WNV infections in five North American raptor species, for which no clinical signs were observed (Komar et al. 2003; Nemeth et al. 2006a).
In the current study, mosquito salivary glands extracts from Culex pipiens, considered the main WNV vector, were used in the subcutaneous inoculation because of their proven enhancement of early WNV infection in vertebrate hosts (Syter et al. 2006, 2011; Schneider et al. 2006; Schneider and Higgs 2008). Despite that, no clinical signs were observed. Many explanations could be evoked, such as viral dose, but our aim was to mimic a natural infection rather than compare it with a needle challenge. In addition, the clearance of the virus and the rapid appearance of neutralizing antibodies could explain the lack of clinical signs seen in our experimental infection, as previously hypothesized (Nemeth et al. 2006a). On the other hand, the high viral load and the microscopic lesions observed in the CNS at 14 dpi may indicate that neurological clinical signs could have developed later. Although we report a subclinical WNV infection in Gyr-Saker hybrid falcons, the pathological lesions and virus spreading in tissues observed here might have an unfavorable impact on the wild falcon population living in its natural environment in terms of morbidity and mortality.
The falcons used in this study were a cross-breed of Ɛ Gyr falcon and ¼ Cherrug and it should be acknowledged that genetic differences between the parent strains might have influenced the results. However, given that all falcons responded similarly to infection, including tissue dissemination, it is hardly arguable that a single dominant gene exists to completely control infection. However, definitive conclusions cannot be drawn from this study due in part to the low number of experimental birds and lack of data about the parental strains. Nonetheless, the falcons used here are a particular cross-breed that might differ from natural falcon populations and species in their susceptibility to WNV infection.
The non-suppurative encephalitis and myocarditis as well as some of the other microscopic lesions observed in the infected falcons were similar to those previously reported in naturally-infected (Fitzgerald et al. 2003; Wunschmann et al. 2004,2005; Nemeth et al. 2006a,2006b; Saito et al. 2007; Lopes et al. 2007; Ellis et al. 2007; Erdelyi et al. 2007; Wodak et al. 2011) and experimentally-infected (Nemeth et al. 2006a) North American raptors. Surprisingly, the scarce amount of viral antigen detected by immunohistochemistry does not correlate with the WNV-RNA load detected by RT-qPCR in the same tissues. One hypothesis to explain this fact, apart from the different sensitivity of these techniques, would be that the lesions could have been induced by the immunological response rather than by viral replication. This hypothesis is supported by the finding of an evident inflammatory reaction in the CNS of the falcon euthanized at 14 dpi. This mechanism has been hypothesized for acute necrotizing encephalopathy of humans associated with influenza virus (Ito et al. 1999).
It is recognized that one TCID50 is equivalent to approximately 0.7 pfu (Dulbecco et al. 1988). Therefore the WNV serum titters from 3–6 dpi of 104 TCID50/mL could be estimated to correspond to 103.8 pfu/mL. This value would be below the threshold level of infectious viremia of WNV, which is estimated at 105 pfu/mL (Komar et al. 2003). These viremia levels would indicate that WNV-infected Gyr-Saker hybrid falcons would not be considered a good reservoir for WNV. However, further experiments of vector competence should be carried out to evaluate the capability of different mosquito species to transmit WNV, and to assess whether such viremias from WNV-infected falcons could be infective for selected mosquito species.
Viral shedding was detected from oral, cloacal, and ocular swabs in all the infected falcons. As WNV was mainly isolated from oral swabs and feather pulps, these samples would be useful for WNV detection, but also could potentially be a source of WNV via oral transmission, as previously reported (Banet-Noach et al. 2003; Docherty et al. 2004; Nemeth et al. 2009). Transmission of WNV via direct contact has been demonstrated in several wild bird species, but not in American kestrels (Falco sparverius) (Komar et al. 2003). However, given the shedding of virus in Gyr-Saker hybrid falcons and their close contact with humans at falconries and wildlife rehabilitation centers, care should be taken when handling these birds until further transmission studies are carried out.
In summary, the results obtained in the present study demonstrate that although Gyr-Saker hybrid falcons do not seem to be a good reservoir for WNV transmission via mosquito, they can become infected with WNV, develop viremia and antibodies, and are able to shed the virus.
Footnotes
Acknowledgments
This work was financially supported by the EU-NADIR project. The WNV NY-99 strain was kindly provided by Dr. Joan Carlos Saiz (CISA-INIA). We would like to thank BSL-3 staff from CReSA for their excellent management and technical assistance. We also thank to Dr. Ursula Höfle for her immunohistochemistry advice, and to Dr. Sebastian Napp for his revisions and suggestions.
Author Disclosure Statement
No competing financial interests exist.