
Abstract
Bartonella are hemoparasites exploiting a range of mammals as reservoir hosts. Several species are zoonotic pathogens. Fleas, lice, and other arthropods, such as ticks, have been implicated as vectors. While the competence of ticks as vectors of Bartonella species has recently been demonstrated, the epidemiological significance of ticks as vectors of Bartonella species in wildlife populations remains unknown. We used the presence of deer at study sites to control the presence of Ixodes ricinus ticks, and used this system to determine whether I. ricinus contributes to the epidemiology of Bartonella species infections in small mammals. Ticks were present at all sites with deer, but were absent from all sites without deer; however, the abundance of ticks on small mammals did not affect the probability of wood mice being infected with Bartonella species. Data presented here indicate that I. ricinus is not involved in the transmission of Bartonella in woodland rodents.
Introduction
The current study investigates the relative importance of ticks as vectors for small mammal-associated Bartonella spp. under natural conditions. This was achieved by comparing the epidemiology of Bartonella infections in tick-infested and tick-free small mammal populations across Northern Ireland. It was predicted that if dependent on ticks for disease transmission, the prevalence of Bartonella spp. in small mammals will be lower where ticks are absent in comparison to where ticks are present. We make use of the strong positive relationship between ticks and wild deer and the patchy distribution of the latter to make comparisons of disease status in the European wood mouse Apodemus sylvaticus.
Materials and Methods
Sampling of wood mice, Apodemus sylvaticus
Ten mixed broadleaf and coniferous woodland sites were sampled throughout Northern Ireland, United Kingdom, over 8 weeks in May, June, and July 2007 (Fig. 1). Five sites did not have large mammals present, while the remaining five had resident populations of fallow deer, Dama dama (3 sites), and red deer, Cervus elaphus (2 sites), as determined by historical records, local expert knowledge, and field surveys. One-hundred and eighty self-set snap traps were set in pairs, 15 m apart in vegetation next to forest tracks. The traps were set after 6

Locations of study sites in Northern Ireland; solid circles represent sites with deer and ticks present, and open circles represent sites without deer and ticks.
Survey of questing ticks
Questing ticks were surveyed using a standardized drag sampling technique. A 1×1-m piece of towelled material, weighted and spread out with bars at the leading and rear edges, was dragged at 1 m/sec−1 along vegetation adjacent to forest tracks. The drag was inspected after each 15-m transect with a total of 20 transects per site. In addition to standardized methods, additional non-standardized drags were also conducted to increase the sample size available for pathogen detection. Ticks were removed using fine forceps, and stored in 70% ethanol for processing as described. Questing ticks were surveyed at the same time as wood mice, and only on dry sunny days.
PCR detection of Bartonella species
Crude DNA extracts were prepared from blood samples and ticks as previously described (Bown et al. 2003). To control for cross-contamination, a negative control (water only) was processed concurrently with every five samples. DNA from Bartonella species was detected using a semi-nested genus-specific PCR assay targeting a fragment of the 16S-23S rDNA intergenic spacer region (ISR) (Telfer et al. 2005), and a nested genus-specific PCR assay targeting a gltA fragment (Norman et al. 1995). PCR success was determined by electrophoretic resolution of amplification products on agarose gels stained with ethidium bromide, followed by examination under UV light. The preparation of all PCRs was carried out in purpose-built laboratories, and templates for first and second round assays were added in separate, dedicated rooms. Downstream processing of amplification products was carried out in a laboratory remote from those used for the preparation and execution of PCRs. In addition to the cross-contamination controls mentioned, each PCR also included a reagent control (no DNA) and a positive control (B. bacilliformis DNA). Selected amplification products were purified using a Qiaquick kit (Qiagen Inc., Valencia, CA), and sequenced in both directions by a commercial sequencing service (Macrogen, Seoul, Korea) using the same primers as those used for amplification. Sequence data were assembled, verified, and analyzed using BioEdit v7.0.9© (Ibis Biosciences, Carlsbad, CA).
Statistical analysis
Generalized linear mixed models (GLMMs) were used to investigate whether the abundance of I. ricinus ticks on wood mice affected the probability of an individual wood mouse being infected with Bartonella spp. The variables tick load, sampling date, sex, age, and mass, and 2-way interactions for sex, age, and mass were included in the GLMMs, with the presence or absence of Bartonella spp. as a binomial response variable, a logit link function, and site as a random effect (to account for between-site variations). In order to remove the confounding effects of pregnancy (via body mass and immunity) on the probability of infection, pregnant females were removed from the analyses. All model permutations were created and the Akaike Information Criterion (AIC) calculated for each model. Models within 2 AIC units (Δ
i
<2) of the top model were retained as a top set of models, and the Akaike weight (0 ≤wi
≤1), a measure of the strength of evidence for each model relative to other models, was calculated for each model in the top set (Burnham and Anderson, 2002). All analyses were conducted using the lmer procedure within lme4 in the R software package available under GNU license at
Results
Abundance of wood mice and ticks
Two-hundred eighty-eight wood mice were trapped over 18 nights with a mean of 29.60±3.36 (SE) mice caught at deer sites, 28.0±3.2 at no deer sites, and 28.8±2.2 caught per site overall (Table 1). In total, 1168 ticks, all identified as I. ricinus, were recovered from wood mice; 1165 (99.7%) were larvae and three (0.3%) were nymphs. The presence of ticks was dependent on the presence of deer at each site. Thus ticks were completely absent from mice in forests without large mammals, but were present on mice at all sites with deer present. Overall mean larval and nymphal tick loads on mice at sites with deer were 7.87±1.09 and 0.02±0.01, (SE) respectively. Two-hundred and thirty-three questing ticks (100 larvae, 129 nymphs, and 4 adults) were collected from standardized drag samples, and all were identified as I. ricinus. Again, ticks were found at every site with deer present, but were not present at any sites without deer (Table 2). The density of ticks on vegetation was low, with numbers/m2±SE of 0.086±0.019 for larvae, 0.067±0.014 for nymphs, and 0.003±0.001 for adults. An additional 167 nymphs and 6 adults were collected by non-standardized drags. Only I. ricinus was recovered from these non-standardized drags, and only at sites with deer present.
Prevalence and diversity of Bartonella species
DNA extracts were prepared from the blood of 270 wood mice and 277 questing ticks (267 nymphs and 10 adults), and incorporated into Bartonella genus-specific PCRs. Bartonella DNA was detected in 81 (30.0%) wood mice, with the prevalence of Bartonella DNA at individual sites varying between 3.0% and 80.6% (Fig. 2). Six (2.2%) ticks (five nymphs and one adult) were found to contain Bartonella DNA. Sequence data for a 338-base pair gltA fragment were obtained for Bartonella present in 16 mice and one adult questing tick. Comparison of these sequences indicated that all shared > 96% similarity with the gltA sequences of either B. taylorii or B. birtlesii, and <92% similarity with the gltA sequences of other members of the genus. Sixteen sequences (from 15 mice and the tick) shared between 90.5% and 98.2% similarity with that of B. taylorii type strain (M6), whereas one sequence (from a mouse) shared 99.7% similarity with that of the B. birtlesii type strain (IBS325). The gltA sequences obtained in this study have been deposited in GenBank and have been allocated the following accession numbers: JN228372–JN228375.
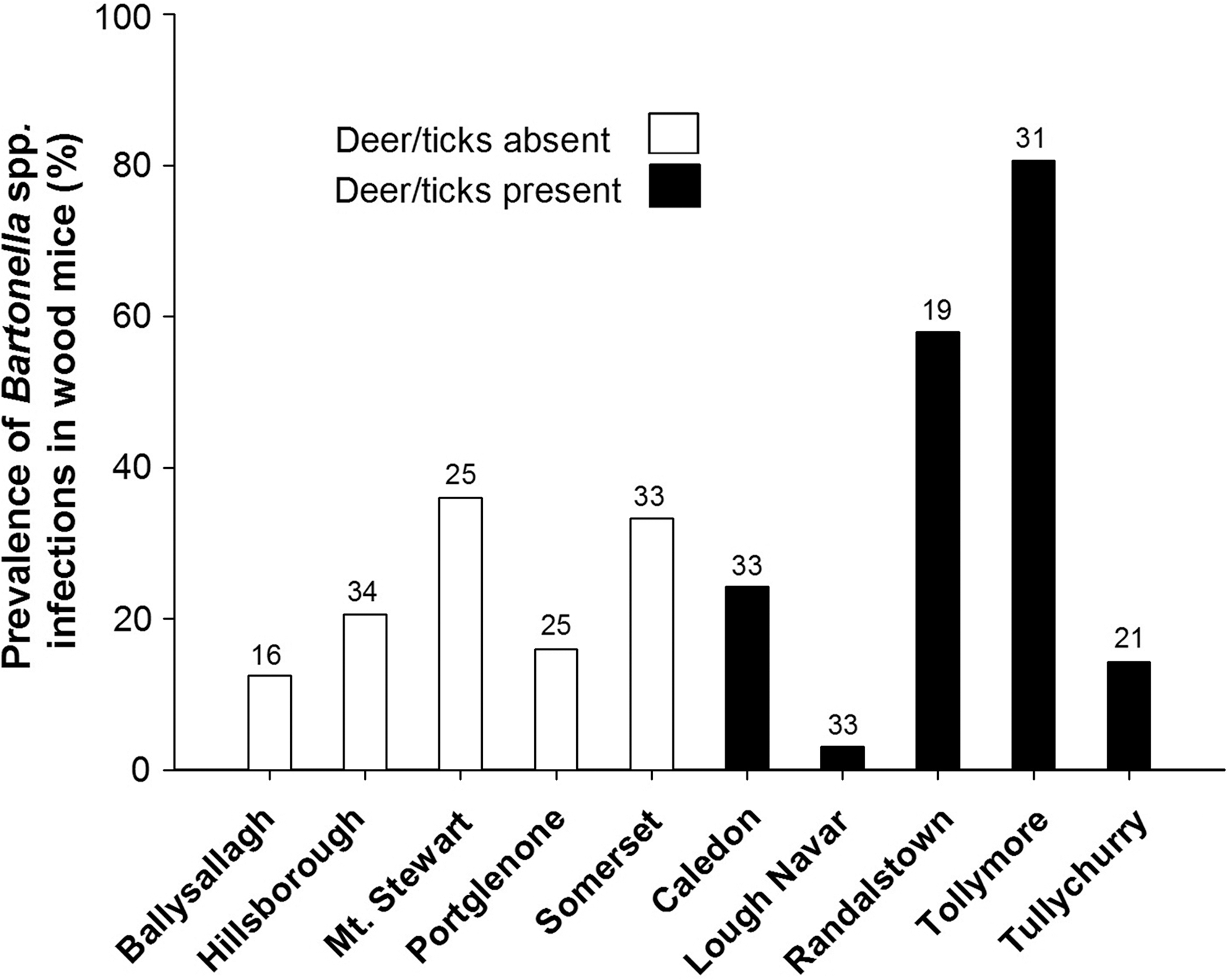
Prevalence of Bartonella spp. infections in the wood mouse Apodemus sylvaticus at sites without deer and ticks, and sites with deer and ticks present, across Northern Ireland. The figures above the bars indicate the number of samples screened for Bartonella spp. infections at that site.
Factors affecting the probability of Bartonella species Infection
Six top models were selected according to AIC (Table 3) from GLMMs investigating factors affecting the probability of an individual wood mouse being infected with a Bartonella species. The variable “sex” was significant (p <0.001) in every model within the top set, and was the only variable present in the single top model, with males more likely to be infected (probability of infection=0.35) than females (0.15). The variable “tick load” was present in a single model within the top set, but had no significant effect on the probability of infection. The mean prevalence of infection of Bartonella spp. in mice at sites without deer present was 23.7±4.7 (%±SE), while the mean prevalence of infection at sites with deer present was 36.0±14.4.
The Akaike weight of each model (0 ≤wi ≤1) and corresponding p values for variables within the each model in the top set are presented. Single top models, and variables significant at the p <0.05 level are in bold.
Discussion
We showed that the presence or absence of ticks had no effect on the probability of infection of wood mice with Bartonella spp., and thus demonstrated that I. ricinus did not contribute significantly to cycles of infection in wood mice in our study area. Therefore, there must be one or more other arthropod vectors maintaining infections of Bartonella spp. in wood mouse populations.
Our findings are in accordance with those reported in a study based in Cantwell, Alaska, where 14% of northern red-backed voles (Myodes rutilus) were infected with a Bartonella species, despite ticks being completely absent from the area (Matsumoto et al. 2010). The Bartonella recovered in the study were thought to be the same “Grahamella-like organism” that Fay and Rausch (1969) had previously used to infect M. rutilus by inoculating them with the homogenate of Megabothris abantis, thus implicating this flea in their transmission. The two species of Bartonella identified in the current study, B. birtlesii and B. taylorii, are adapted to woodland rodents (Bown et al. 2004; Telfer et al. 2005) and have been detected previously in wood mice from the U.K. and Ireland (Birtles et al. 2001; Telfer et al. 2005). Experimental data have shown that the flea, Ctenophthalmus nobilis, is a competent vector of B. grahamii and B. taylorii in bank voles, Myodes glareolus (Bown et al. 2004). Unfortunately, data on fleas were not available in the current study, as fleas leave the host posthumously (Cole and Koepke 1947; Gross and Bonnet 1949; Stark and Kinney 1962), and snap-trapping was the method we utilized to sample wood mice. However, C. nobilis is the most common flea found on wood mice in Ireland (Langley and Fairley 1982; Telfer et al. 2005), and fleas containing Bartonella DNA have been collected from wood mice that were also bacteremic with Bartonella spp. (Telfer et al. 2005). Therefore, it is highly probable that C. nobilis is the vector of B. birtlesii and B. taylorii in free-ranging wood mouse populations in Ireland, and as we have demonstrated, that I. ricinus does not significantly contribute to their transmission.
It is noteworthy that we, like others have done previously (reviewed by Angelakis et al. 2010), demonstrated the presence of Bartonella DNA in a small proportion of the questing ticks we surveyed. This observation, particularly in the context of the conclusions of our study, re-emphasizes the often-expressed view that the mere presence of Bartonella DNA in ticks does not prove vector competence or confer epidemiologic significance (Billeter et al. 2008; Telford and Wormser 2010). Furthermore, although a very recent study has reported experimental transmission of B. birtlesii from infected to uninfected laboratory mice by I. ricinus ticks (Reis et al. 2011), thereby demonstrating vector competence, our findings bear out the caveat expressed by others (Telford and Wormser 2010), that even with this demonstration, additional data are needed to conclude that Ixodes spp. are epidemiologically relevant as vectors of Bartonella spp. in natural populations.
The irrelevance of ticks for rodent-associated Bartonella is further supported by the feeding behavior of I. ricinus. For trans-stadially transmitted tick-borne pathogen cycles to develop, more than one tick life stage must acquire a blood meal from a given host species (Randolph and Storey 1999). In Ireland, the abundance of nymphs feeding on small mammals is low, with a ratio of 1 nymph to 388 larvae (Harrison et al. 2011). Therefore, even if I. ricinus was a competent vector, tick-borne cycles of Bartonella spp. may find it difficult to become established, or persist, in small mammal populations given the scarcity of non-larval tick life stages on wood mice.
This is the first study to observe a male sex bias in the prevalence of infection of Bartonella spp. in wood mice. Male sex-biased parasitism occurs across a wide range of mammalian taxa (Schalk and Forbes 1997; Moore and Wilson 2002), and a male sex bias in parasitism rates by I. ricinus has been observed in wood mice in Ireland (Langley and Fairley 1982; Harrison et al. 2010). Telfer and associates (2005) also found a male sex bias in the parasitism rates of C. nobilis infesting bank voles in Ireland, but did not find any sex bias for sympatric wood mice. The reason for male sex-biased parasitism is unclear and is widely debated in the literature (Schalk and Forbes 1997; Moore and Wilson 2002; Harrison et al. 2010). In the case of Bartonella spp. infections, males may be more heavily infected simply because they are infested with more arthropod vectors than females, or alternatively, they may be more prone to infection by the hemoparasites themselves. For example, sex differences in behaviors such as grooming or mate seeking will affect the number of arthropod vectors present on a host (Mooring et al. 2004). Males may also suffer from the immunodepressive effects of testosterone (Hamilton and Zuk 1982; Folstad and Karter 1992). Wood mice are sexually size dimorphic, and therefore body size may also contribute. Larger males may provide a larger surface area for ectoparasite vectors (Shine 1989) or, under limited resources, males may expend energy growing large at the expense of their own immunity (Rolff 2002).
Results of the current study suggest that I. ricinus does not contribute to the epidemiology of Bartonella infections in wood mouse populations in Ireland, and that infections with this pathogen are maintained by another arthropod vector or vectors, most likely fleas.
Footnotes
Acknowledgements
A. Harrison was supported by a Ph.D. studentship from the Department of Agriculture and Rural Development (DARD), and access to field sites was kindly provided by the Forest Service of Northern Ireland. This study was conducted in compliance with the ethical procedures of the Queen's University of Belfast.
Author Disclosure Statement
No competing financial interests exist