
Abstract
As a response to the need for improved and cost-efficient West Nile virus (WNV) and other flavivirus surveillance tools, we tested 887 juvenile free-living red deer, 742 free-living juvenile wild boar, and 327 farmed deer, to detect temporal variability in exposure to these viruses. Thirty of 742 wild boar samples (4%; 95% CI 2.8,5.7) yielded a positive ELISA result. Antibody-positive individuals had been sampled between 2003 and 2011 in localities from central and southern Spain. No wild boar from the northern half of Spain (n=120) tested positive. Regarding juvenile wild red deer, only two out of 887 samples yielded a positive ELISA result (0.2%; 95% CI 0.1,0.8). These two samples came from the same site and sampling year. The likelihood of detecting contact with WNV or cross-reacting flaviviruses was 18 times higher among juvenile wild boar than among juvenile red deer. ELISA positivity among farmed deer increased 10-fold after local flavivirus outbreaks recorded in the summer and autumn of 2010. This survey demonstrated the potential usefulness of juvenile wild ungulates, particularly wild boar, as suitable flavivirus sentinels in southwestern Europe, and that systematic serum banking of samples from hunter-harvested wildlife or from individual farmed ungulates provides valuable material for retrospective epidemiological surveys and future disease monitoring.
Introduction
Antibodies against WNV have been reported in several bird species, with seroprevalences ranging from 2–43% (Figuerola et al. 2007,2008); thus WNV is probably endemic and widespread in its usual bird-mosquito cycle in Mediterranean Iberia (Hayes et al. 2005). However, few human cases of WN fever have been recorded (Bofill et al. 2006; Kaptoul et al. 2007;
Hunter-harvested mammals often are more accessible than wild birds and allow sampling larger amounts of serum. Thus it has often been suggested to use mammals as substitute flavivirus sentinels because contact detection would indicate transmission outside the enzootic bird cycle (Root et al. 2005; Teehee et al. 2005; Gómez et al. 2008; Blitvich et al. 2009). However, medium-sized and large mammals generally have a long life span, and antibodies against WNV may persist more than 1 year (Geevarghese et al. 1994). Hence, the annual cohorts of juvenile mammals (after losing their maternal antibodies) are potentially the best indicators of current flavivirus circulation. Studies in white-tailed deer (Odocoileus virginianus) demonstrated that it is unlikely that they are an important amplifying host for WNV (Farajollahi et al. 2004), but that clinical disease and mortality are possible (one fatal case was described by Miller and associates [2005]). Several authors demonstrated seroconversion with enzyme-linked immunosorbent assays (ELISA), and by plaque-reduction neutralization tests (PNRT), and prevalences obtained ranged 0.9–12.7% (Farajollahi et al. 2004; Santaella et al. 2005). Regarding the Eurasian wild boar (Sus scrofa), we assume an analogous response to that of domestic pigs (Sus scrofa). Pigs develop low viremias of short duration, and it is unlikely that they are amplifying hosts, but due to their serological reaction, pigs can be useful as sentinels (Teehee et al. 2005). Also, flavivirus antibody prevalences have been reported in wild boar from the Czech Republic (6.5%; Halouzka et al. 2008), and feral swine in the U.S. (22.5%; Gibbs et al. 2006). A preliminary exploratory study of Eurasian wild boar from Spain revealed exposure of the species to WNV in areas with high reported WNV activity in birds (Gutiérrez-Guzmán et al. submitted). Cattle would be easier to sample than wild ungulates. In fact, a 4% seropositivity was recorded in Turkey (Ozkul et al. 2006). However, one study in Spain found no seropositive cattle in a wetland where WNV was known to circulate among birds and equids (Jiménez-Clavero et al. 2007). We hypothesized that in the Iberian Peninsula, red deer (Cervus elaphus) and Eurasian wild boar would come into contact with members of the Flaviviridae family at similar rates as sympatric wild birds, and that subsequent seroconversion would be easier to monitor since these abundant and widespread game species exhibit several advantages compared to birds: (1) a relatively long lifespan, (2) a limited home range and less mobility than their avian counterparts, (3) easy and cost-effective sampling of hunter-harvested animals and in game farms, and (4) larger quantities of serum available. With the aim of adding a useful tool to the WNV and flavivirus surveillance toolbox, we tested a large number of juvenile red deer and wild boar to detect temporal variability in exposure to WNV or cross-reacting members of the genus Flavivirus.
Materials and Methods
Sampling
Serum samples were collected between 2000 and 2011 from hunter-harvested Eurasian wild boar (n=742) and red deer (n=862). Samples were stored frozen at −20°C until analysis. Only wild boar between 4 and 12 months of age were studied. Regarding deer, the animals included in this part of the study were between 4 and 18 months old.
In addition, we had access to individual serum samples obtained from 327 farmed red deer of all ages except calves in Cádiz, southern Spain. This farm (indicated by an asterisk in Fig. 1) is located in the region where a WNV (
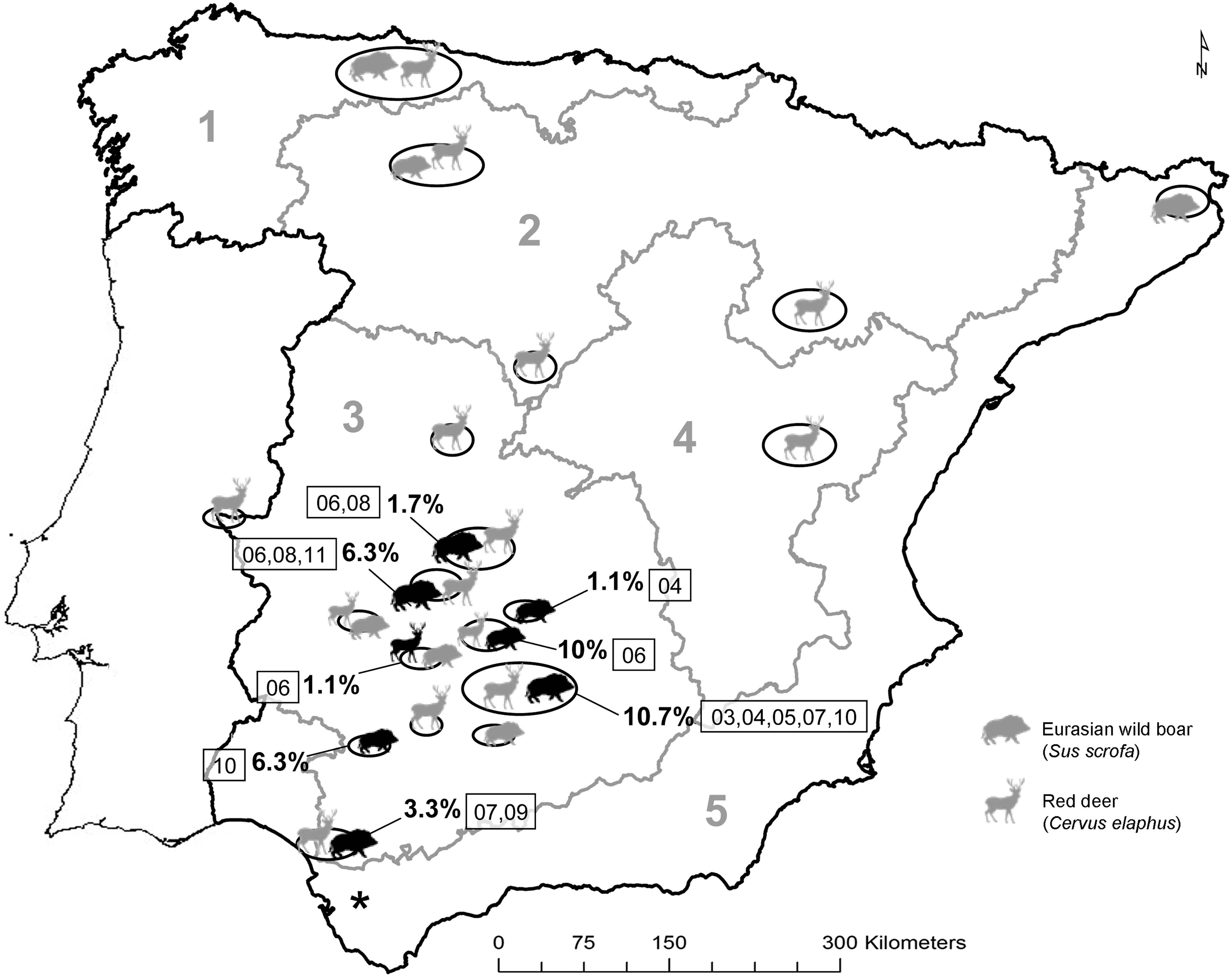
Map of the Iberian Peninsula showing the 19 sample sites and their flavivirus antibody seroprevalence. Different shapes represent the sampled species, red deer (Cervus elaphus) and wild boar (Sus scrofa) and their sizes are proportional to the sample (large symbols are for sample sizes greater than 50 animals, while small symbols are for sample sizes below 50 individuals). Grey shapes indicate negative results and black shapes positivity. Mean seroprevalences are indicated by percentages. The years when seropositive animals were detected are the numbers in the squares. The asterisk marks the location of the red deer farm in Cádiz. Grey numbers indicate the five peninsular Spanish Bioregions.
ELISA test
A WNV blocking ELISA was used to screen for flavivirus-specific immunoglobulin G (IgG) antibodies (Ingenzim WN Compac®; Ingenasa, Madrid, Spain) (Sotelo et al. 2011). Following the manufacturer's instructions, samples with percent of inhibition >40% were considered positive. Although the assay uses the protein E of WNV, this method may detect cross-reactive antibodies against antigenically-related flaviviruses of the Japanese encephalitis complex. Due to logistical limitations positive sera could not be confirmed by seroneutralization.
Statistical analysis
Comparison of seroprevalences between species and sampling periods was performed by means of chi-square tests. Only wild boar from sites with n>10 in both time periods were considered for the time analysis. Generalized linear models (GLM) with binomial distribution and logit link function were used for calculating the effect of sampling year (continuous), the sampling area (categorical, n=6), and their interaction, on the probability of testing positive on the ELISA (binomial, 1 positive, 0 negative). Areas without stratified sampling per year were excluded from the analysis. We used a backward stepwise strategy to obtain the final model, selected by the best p value. Data were analyzed using the IBM SPSS statistical package, version 19.0 (IBM Corporation, Somers, NY).
Results
As shown in Table 1, only 30 of 742 juvenile wild boar samples (4%; 95% CI 2.8,5.7) yielded a positive ELISA result. These antibody-positive individuals had been sampled between 2003 and 2011 in localities from central and southern Spain (Bioregion 3), where sample sizes were larger (Table 1 and Fig. 1). No wild boar from the northern half of Spain (n=120) tested positive. When analyzing over time periods (n=612), wild boar sampled between 2000 and 2005 showed a seroprevalence of 7%, and in those sampled between 2006 and 2011 the seroprevalence remained stable at 3.4% (χ2=3.2, 1 d.f., p>0.05). The best model was explained by the area of sampling (Wald χ2=12.2, 2 d.f., p<0.05), while year was not a significant factor in the model. Modeling revealed no significant time trend (β=0.067, p>0.05).
The 327 farmed red deer samples were from Bioregion 5.
Regarding juvenile wild red deer, only two out of 887 samples yielded a positive ELISA result (0.2%; 95% CI 0.1,0.8; Table 1). These two samples came from the same site (a private hunting estate in south-central Spain), and had been collected in January and December 2006 (Fig. 1). No wild boar samples were available for 2006 for this site. The likelihood of detecting contact with WNV or cross-reacting flaviviruses was 18 times higher among juvenile wild boar than among juvenile red deer (χ2=30, 1 d.f., p<0.001).
Figure 2 presents the results obtained from the individually surveyed farmed red deer. ELISA positivity among deer increased 10-fold after the lineage 1 WNV outbreak that affected horses, and the parallel BagV outbreak that affected game birds in the same locality. Overall seroprevalences in winter 2008–2009 and winter 2009–2010 were 1.2% and 3%, respectively. Samples collected in January 2011 from the same individuals yielded 38.8% seropositivity. Among these 327 red deer sera, two tested positive in 2009, negative in 2010, and positive again in 2011, and 6 tested negative in 2009, positive in 2010, and negative again in 2011. Two deer consistently tested positive in all three years. All other sera either tested consistently negative (58.7%) or tested positive only in January 2011 (36%).
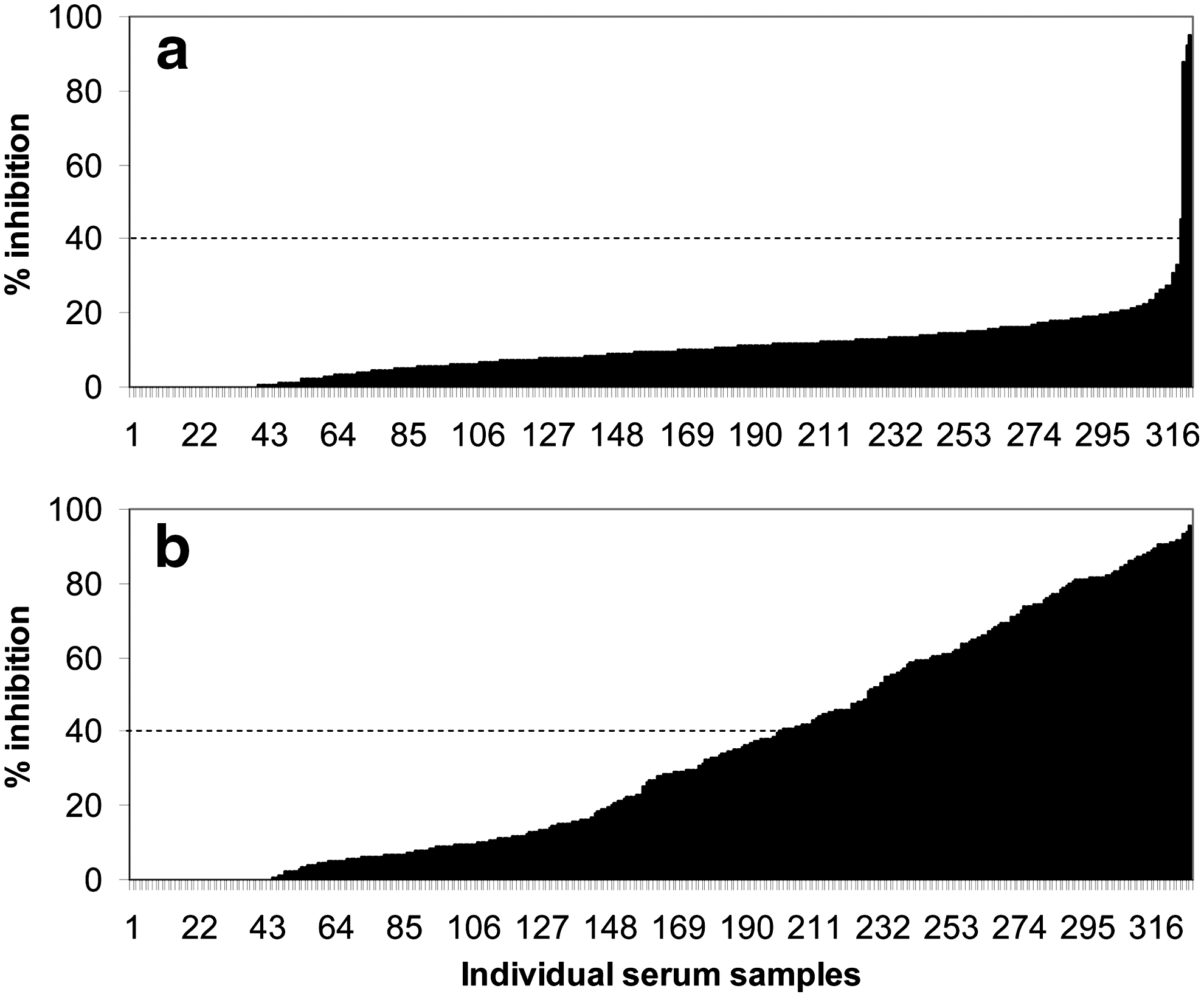
Individual serum antibody levels (expressed as percent inhibition) against West Nile virus or cross-reacting flaviviruses detected by ELISA in 327 farmed red deer from Cádiz, southern Spain. Data are presented for December 2008/January 2009 (
Discussion
This survey demonstrated the potential usefulness of wild boar, and to a lesser extent of red deer, as sentinels for WNV or cross-reacting members of the genus Flavivirus. It also showed the value of routine serum banking for retrospective epidemiology to provide an easy and cost-effective way to monitor the current year activity of a given pathogen (flavivirus) in a region (Boadella et al. 2011a).
Based on the literature, we expected similar seropositivity rates in deer and wild boar (Santaella et al. 2005; Halouzka et al. 2008). However, free-living juvenile wild boar appeared to be far more suitable as sentinels than free-living juvenile red deer. Several not mutually exclusive hypotheses might explain the differences in antibody prevalence observed between free-living juvenile wild boar and red deer. First, the low hair density, the low thickness of the epidermis, and other peculiarities of wild boar skin (Meyer et al. 2011) make them more susceptible to mosquito bites than deer; second, their scavenging behavior, as WNV (and possibly other flaviviruses) can occasionally be transmitted orally via consumption of infected prey or carrion as previously demonstrated in mammals (Austgen et al. 2004); third, wild boar (and pigs) might be more likely to produce an antibody response than ruminants, as is observed for tuberculosis (Boadella et al. 2011b); and fourth, since host preferences are common factors modulating vector-borne diseases (Zwiebel and Takken 2004), we suggest that flavivirus vectors may display a preference for suids.
The only two seropositive wild red deer were recorded in the same site and during the same year. This suggests an epidemiological link between both events, rather than coincidence, and shows that using juveniles for surveillance purposes can be effective. Unfortunately, no wild boar were sampled in this site the same year, precluding any comparisons. However, the higher apparent sensitivity of wild boar as flavivirus sentinels allowed detection of antibodies in all years since 2003 (sample sizes were low before 2003), and the analysis over time showed a stable or even declining seroprevalence. This would confirm that WNV (or cross-reacting flaviviruses) have been circulating for years in the Iberian Peninsula, as reported for birds of prey (Höfle et al. 2008), aquatic birds (Figuerola et al. 2007), and for humans (Garea Gonzalez and Filipe 1977; Lozano and Filipe 1998). A very important aspect of WNV surveillance is early detection (Rockx et al. 2006). The results reported herein prove that antibodies can be detected in wild ungulates, even before cases are detected in horses or humans, for instance in central Spain.
The fact that two of the farmed deer consistently tested positive in 2009–2011 could suggest that antibodies against flavivirus may persist for periods over 1 year, or alternatively it could suggest re-exposure (Geevarghese et al. 1994). This confirms the choice of juveniles for flavivirus surveillance, since antibodies in these animals would indicate a recent exposure to the agent. This study also demonstrated that farmed deer offer an easily accessible sample that can be used to detect seroconversion. Finally, the fact that neither deer nor wild boar from the northern third of the peninsula had a positive ELISA result may be due to two facts. First, contact with flavivirus is not a frequent event among these wild ungulates, or second, the lower sample sizes compared to Bioregion 3, and thus a reduced detection probability.
In addition to wild and domestic birds, horses would seem the most straightforward sentinel species for WNV and related flaviviruses (Jiménez-Clavero et al. 2007). However, well maintained horses are often vaccinated, impeding their use as sentinels, and access to semi-free-ranging unvaccinated horses would seem as difficult to access to hunter-harvested wild ungulates. Regarding wild ungulates, sampling could eventually be further facilitated by using soft tissue extracts (e.g., lung extracts) as an alternative to serum (Ferroglio et al. 2000).
In conclusion, we confirmed that juvenile wild ungulates, particularly wild boar, are suitable flavivirus sentinels in southwestern Europe, and that systematic serum banking of samples from hunter harvested wildlife or from individual farmed ungulates provides valuable material for retrospective epidemiological surveys and future disease monitoring.
Footnotes
Acknowledgments
This study was supported by POII09-0141-8176 (Castilla-La Mancha) and by JCCM PPIC10-0226-0243. The authors also acknowledge the support of the Ministerio de Medio Ambiente, Rural y Marino (MARM). This study was performed with the aid of colleagues at IREC that participated in the fieldwork during the entire study period. We also wish to thank J. Queirós for help with the serum database, and to recognize the commitment of J.A. Ortiz to research collaborations.
Author Disclosure Statement
No competing financial interests exist.