
Abstract
Sandfly fever turkey virus (SFTV) is a recently-discovered sandfly fever Sicilian virus (SFSV) variant (family Bunyaviridae, genus Phlebovirus), characterized during retrospective evaluation of febrile disease outbreaks in Turkey. In addition to causing sandfly fever, SFTV was observed to induce elevation of liver enzymes, and to cause thrombocytopenia in affected individuals. This study was conducted to identify vectors for phleboviruses including SFTV in Ankara province, Turkey, where evidence indicates ongoing circulation of SFTV, as well as Toscana virus. Sandfly sampling was performed in Ankara province in the vicinity or in animal housing facilities in 15 peri-domestic sites. Male sandflies were identified morphologically, whereas females were evaluated individually for Phlebovirus RNA via a nested-PCR assay with consensus primers. Selected individuals and PCR-positive sandflies were subjected to barcoding via cytochrome c oxidase sequence analyses. The source of blood meals in virus-infected sandflies was investigated using a multiplexed PCR targeting the mitochondrial cytochrome b gene of various vertebrates. A total of 667 sandflies were captured in 11 locations. Morphological identification of males (n=226) revealed Phlebotomus major sensu lato as the most abundant species (38.9%), followed by Phlebotomus sergenti (20.4%), Phlebotomus halepensis (17.7%), Phlebotomus papatasi (10.2%), Phlebotomus simici (3.98%), Larrousius spp. (3.53%), Phlebotomus tobbi (1.32%), Phlebotomus perfiliewi perfiliewi (1.32%), and others. Virus sequences were detected in 3 (3/441) sandflies, two of which were characterized as P. major s.l. via barcoding. The detected sequences in sandflies were identified as SFTV, and were identical or similar to sequences from patients from the same area and the prototype SFTV strain. Bovine and human blood meals were demonstrated in SFTV-infected sandflies. P. major s.l. has been identified as the vector species for SFTV. Bovidae need to be evaluated as probable amplifying hosts for SFTV.
Introduction
In addition to the SFSV and TOSV, new phlebovirus strains have been identified in sandflies since 2006, including Massilia virus in southern France (Charel et al. 2009), Granada virus in Spain (Collao et al. 2010), Punique virus in Tunisia (Zhioua et al. 2010), Adria virus in Albania (Papa et al. 2011), and novel sequences in Algeria (Moureau et al. 2010). Although serologic data indicate human and/or animal exposure to some of these strains, their pathogenicity has not yet been elucidated, and no association with clinical disease could be established. However, another new SFSV variant, sandfly fever Cyprus virus (SFCV), which has been identified in Greek troops in Cyprus, is clearly associated with outbreaks of febrile disease (Papa et al. 2006).
Sandfly fever turkey virus (SFTV) is a recently-discovered phlebovirus variant that was characterized during retrospective evaluation of outbreaks in 2007–2008 from Ankara (Central Anatolia), Izmir (Aegean coast), and Adana (Mediterranean coast) provinces in Turkey (Carhan et al. 2010). SFTV is provisionally included in the sandfly fever Sicilian serocomplex, and displays 98% and 91.8% nucleotide homology in the S segment to SFCV and SFSV prototype strain Sabin, respectively (Carhan et al. 2010). In addition to the common symptoms of sandfly fever, SFTV infections were frequently aggravated by gastrointestinal symptoms and elevation of liver enzymes, which has also been documented for SFCV (Konstantinou et al. 2007), along with thrombocytopenia (Carhan et al. 2010; Kocak Tufan et al. 2011). A seroprevalence study has demonstrated SFTV exposure in asymptomatic blood donors in Ankara and Konya provinces (Ergunay et al. 2011). Moreover, acute SFTV infections were also identified in Ankara in 2009, which indicates ongoing circulation of the virus in this region (Kocak Tufan et al. 2011).
The activity of various species of phlebotomine sandflies has been observed in the Mediterranean, Aegean, and Central Anatolian regions of Turkey, including P. papatasi and P. perfiliewi, well-known vector species for SFSV and TOSV, respectively (Yagci et al. 1998; Yaman and Dik 2006; Simsek et al. 2007). The aim of this study was to perform a field study to investigate sandflies as phlebovirus vectors in Ankara, where TOSV and SFTV cases have recently been reported (Ergunay et al. 2011; Kocak Tufan et al. 2011).
Materials and Methods
Setting and sample collection
The entomological survey was conducted in Mamak County of Ankara province. Ankara (39°56’N, 32°52’E) is the capital and second most densely-populated city in Turkey, with a population over 4.6 million. It was observed that the majority of SFTV-infected individuals from Ankara resided in the same suburb, the Mamak county, hence the syndrome was informally named “Mamak fever” (Carhan et al. 2010; Kocak Tufan et al. 2011). Mamak (478 km2, elevation 899 m) is one of the largest rural counties of Ankara, with over 503,000 inhabitants in the northeastern part of the city.
For the sandfly sampling in or near Mamak County, 15 locations were selected. Sampling was performed for three consecutive days each week during July 2011. CDC miniature light traps (John W. Hock Company, Gainesville, FL) equipped with an ultra-fine mesh were employed in each station. The traps were placed 1–2 meters above ground in the vicinity or in animal housing facilities (cow barns) in peri-domestic sites, and kept on site from late afternoon until dawn. Captured sandflies were collected in the morning, placed individually into 1.5-mL microcentrifuge tubes according to gender and trapping location, and transferred to the laboratory on ice. The head and genitalia of male sandflies were dissected and visualized on slides prepared with Swan solution, and were examined for morphological identification to species level via published keys (Theodor 1958; Artemiev 1980; Lewis 1982). The remaining body parts were stored in absolute ethanol until DNA extraction. Females were kept at −80°C for further analysis.
Detection and characterization of phlebovirus sequences in sandflies
Individual female sandflies were homogenized and clarified by centrifugation as described previously (Charel et al. 2009; Zhioua et al. 2010). Two hundred microliters of the supernatant was used for RNA isolation using the High Pure Viral Nucleic Acid Kit (Roche Diagnostics, Mannheim, Germany). Ten microliters from the eluted RNA was used for reverse transcription via random hexamers employing the RevertAid First Strand cDNA Synthesis Kit (Fermentas, Vilnius, Lithuania). All commercial assays were performed according to the manufacturers' recommendations.
For the detection of phlebovirus sequences in individual sandfly homogenates, consensus primers targeting the viral polymerase in the L segment of the viral genome (NPhlebo 1+/1− and 2+/2−) were used in a nested-PCR reaction, as described previously (Sanchez-Seco et al. 2003), with 2.5 U Hot Start Taq DNA polymerase (Bioron GmBH, Munich, Germany), in a PTC-200 Thermal Cycler (MJ Research, Waltham, MA). TOSV ISS.Phl.3 isolate, grown on Vero cells as described previously (Ergunay et al. 2011), was used as a positive control, and extreme care was taken to prevent carry-over contamination. The amplicons were visualized under ultraviolet light after electrophoresis in 2% agarose gels for the expected amplicon size of 244 bases. Detected amplicons from the second round of PCR were cut out and purified with the AXYprep DNA gel extraction kit (Axygen Biosciences, Union City, CA), ligated to the pJET1.2 vector supplied in the CloneJet PCR Cloning Kit (Fermentas), and was used to transform cells as directed by the manufacturer. Forward and reverse primers provided for sequencing were employed for the characterization of cloned amplicons using an ABI Prism 310 Genetic Analyzer (Applied Biosystems, Foster City, CA). The obtained sequences were submitted to GenBank.
Barcoding and blood meal analysis in sandflies
For the characterization of the sandflies, the cytochrome c oxidase I (COI) gene, frequently used for biological barcoding, was employed as described previously (Folmer et al. 1994). A 658-basepair (bp) fragment of the COI gene was amplified via LCO1490 and HCO2198 primers. PCR products were purified using the QIAquick PCR product purification kit (Qiagen, Hilden, Germany), and subjected to cycle sequencing with the amplification primers from both directions.
Phlebovirus PCR-positive sandflies were further evaluated via a multiplexed PCR based on a mitochondrial cytochrome b gene to identify mammalian blood meals employing human, dog, cow, and goat forward, with a universal reverse primer, as previously described (Kent and Norris 2005). The products, visualized in 2% agarose gels, were evaluated for blood meals from humans (334 bp), cows (561 bp), dogs (680 bp), and goats (132 bp), according to amplicon size.
Sequence data analysis
Sequences were edited using BioEdit Sequence Alignment Editor and Chromas (version 2.13) software. The respective sequences and similar sequences available in GenBank were aligned with ClustalW. To construct taxon identity trees, neighbor joining (NJ) and maximum composite likelihood analyses were performed under the assumptions of Kimura's two-parameter model, and bootstrapping of 10,000 data matrices in MEGA 5.0 (Tamura et al. 2011).
Results
Species distribution of phlebotomines
A total of 667 sandflies were captured in 11 of 15 locations (Fig. 1); 226 (33.9%) were male, and 441 (66.1%) were female (Table 1). No trapping was observed in 4 locations despite functioning traps. Morphological identification of the male phlebotomines revealed P. major s. l. as the most abundant species (88/226, 38.9%), followed by P. sergenti (46/226, 20.4%), P. halepensis (40/226, 17.7%), P. papatasi (23/226, 10.2%), P. simici (9/226, 3.98%), Larrousius spp. (8/226, 3.53%), P. tobbi (3/226, 1.32%), P. perfiliewi perfiliewi (3/226, 1.32%), Sergentomyia spp. (2/226, 0.88%), Adlerius spp. (2/226, 0.88%), P. perfiliewi galilaeus (1/226, 0.44%), and Paraphlebotomus spp. (1/226, 0.44%). The numbers and distribution of the sandfly species according to sampling location are provided in Table 1.
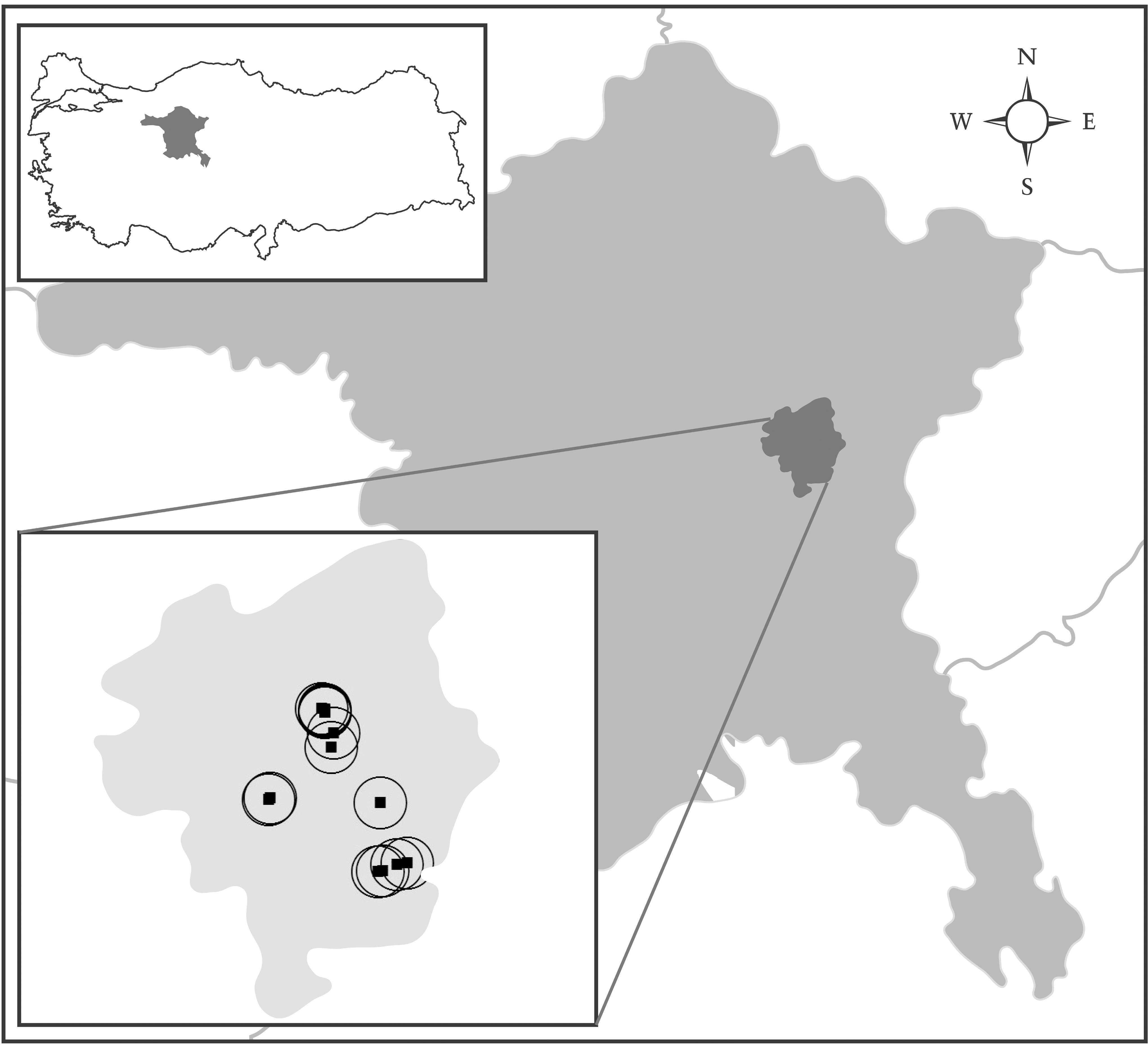
Map of Ankara province illustrating Mamak County and the sandfly sampling sites.
Identification and characterization of phlebovirus sequences in phlebotomines
Three samples (3/441) were observed as reactive in the second round of nested PCR with phlebovirus consensus primers NPhlebo 2+ and 2−. The amplicons were ligated to cloning vector as described above, and 8–10 clones carrying the insert were sequenced. Identical sequences of 204 bp (excluding the primers) were retrieved from all clones analyzed from each sample. BLAST and BLASTX searches in GenBank revealed these sequences to be 98–99% identical to SFTV. Multiple sequence alignment and phylogenetic analysis with similar and divergent phlebovirus sequences from GenBank provided further support for the identification (Fig. 2). Identical sequences were retrieved from sandflies captured at the same location (GenBank accession numbers JN907011 and JN907013; Table 2), which were 1% divergent from the prototype SFTV sequence (GQ847513) in the maximum composite likelihood analysis. The sequence obtained at location A1 (JN907012) was found to be 1% and 2% divergent from the sequences from location AII and prototype SFTV, respectively. To provide evidence for the similarity between SFTV in vectors and human infections, sequences detected in sera of three individuals diagnosed with acute SFTV infections in August 2011 in Ankara were included in the analyses (JN907007, JN907008, and JN907010; Fig. 2). These sequences were obtained via the same methodology used for the phlebotomines, as described above. Maximum composite likelihood analysis revealed identical sequences in one patient and sandflies from location AI, and 98–99% similarity was observed for the remaining sequences (data not shown). No relevant epidemiological connection could be demonstrated between patients and the sampling sites other than being located in the same neighborhood within an area of 5–7 km2.
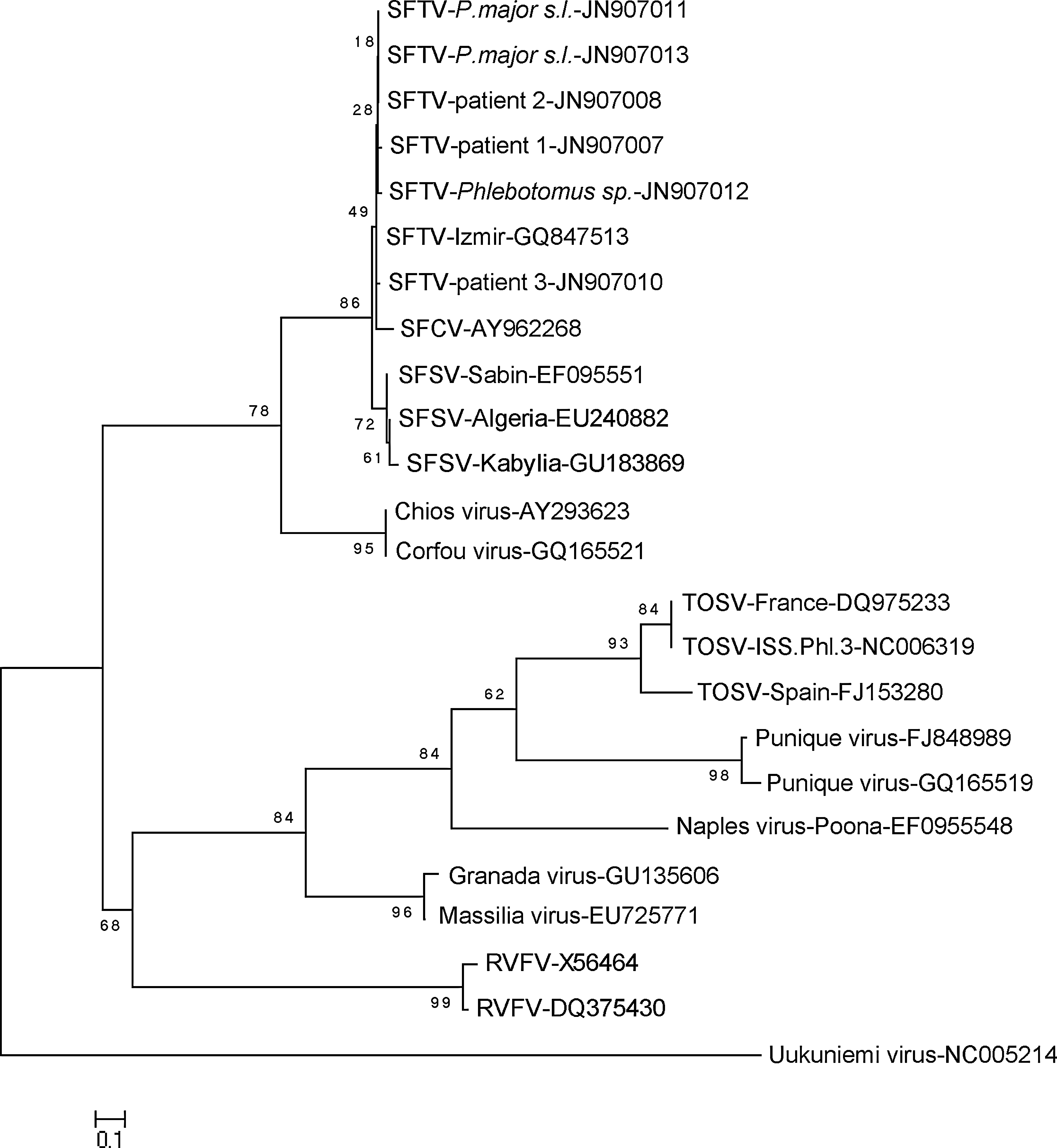
Phylogenetic analysis of the partial sequences of the polymerase gene segment detected in sandflies and patients during July–August 2011 in Ankara province, Turkey. GenBank accession numbers JN907007 to JN909012 represent the sequences included in this study, and GQ847513 is the prototype strain (SFTV, sandfly fever turkey virus; SFCV, sandfly fever Cyprus virus; SFSV, sandfly fever Sicilian virus; TOSV, Toscana virus; RVFV, Rift Valley fever virus).
Molecular characterization of Phlebotomus species detected as phlebovirus vectors
A total of 26 sandflies were captured at the locations (AI and AII) and dates (July 4–6) that included phlebovirus-positive samples (Table 2). Morphological examination of the males (n=13) revealed eight specimens as P.major s.l., four specimens as P. sergenti, and one specimen as P. papatasi. For one female specimen that was reactive in phlebovirus consensus PCR, no amplification could be seen despite repeated tests (Table 2). The COI gene regions of the remaining females (n=12) unidentified morphologically were subjected to sequencing, along with three randomly-selected P. papatasi and P. major s.l. male specimens from different dates. The alignment, including these specimens using the older GenBank entries, and those obtained for several Turkish sandfly species (Erisoz Kasap and Alten, unpublished data) as reference sequences, was 454 bp long. Based on their COI sequences, eight females (including two infected ones) were assigned as P. major s.l., since seven of them were found to have identical sequences with the sympatric P. major s.l. males, and one differed by only five mutations. The NJ analysis also indicated that the rest of the females were clustered with the only P. sergenti sequence (FJ196442.1) available in GenBank, although they fell into two well-supported distinct lineages, which probably reflected intra-species variations (Fig. 3).
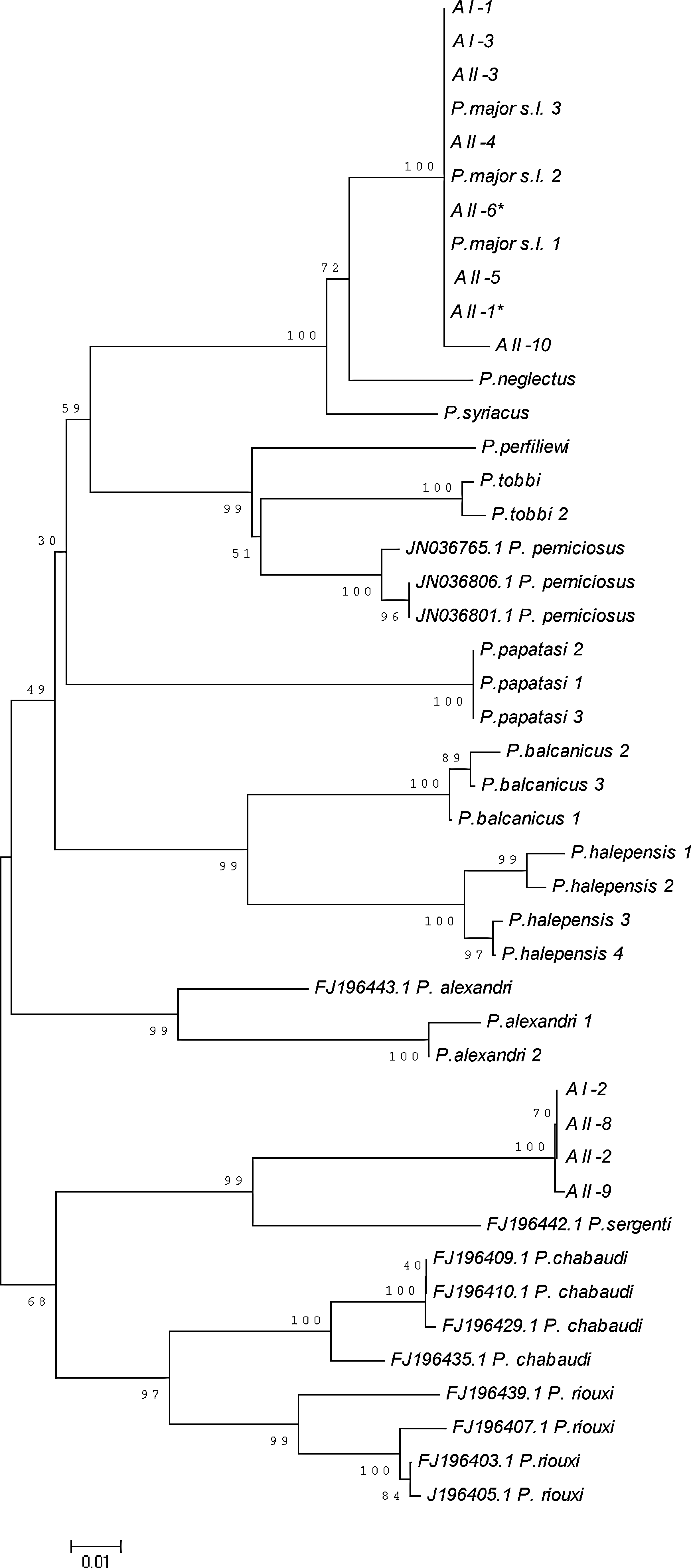
Neighbor joining tree based on the partial cytochrome c oxidase I (COI) sequences used for barcoding of sandflies. AI and AII indicate the sampling locations (see text for details). Phlebovirus consensus PCR-positive samples are indicated with an asterisk.
Blood-fed individuals were observed in 13.2% (58/441) of the female sandflies prior to homogenization, including all phlebovirus-positive specimens. Analysis via multiplexed PCR in these samples revealed bovine blood meals in all and human blood meals in two samples captured in the same location (Table 2).
Discussion
Despite serologic evidence for exposure, detection of acute infections, and documentation of the presence of various species of phlebotomine sandflies, vector surveillance for phleboviruses in Turkey has not been performed previously (Ergunay et al. 2010). Since SFTV is associated with a more severe clinical picture than the typical sandfly fever symptomatology, the identification of phlebotomine species as vectors and their associated habitats are of substantial importance. This study was conducted to identify phleboviruses in sandflies in Ankara province, where SFTV outbreaks were observed from 2007 to 2010, indicating continuous circulation of the virus, in addition to TOSV cases detected in 2010, as well as documented exposure in blood donors (Ergunay et al. 2011, 2011; Kocak Tufan et al. 2011).
Throughout July 2011, we captured 667 sandflies in 11 sampling locations (Table 1). Morphological identification of the male phlebotomines indicated that the most frequently-observed species were P. major s.l. (38.9%), P. sergenti (20.4%), P. halepensis (17.7%), and P. papatasi (10.2%), followed by others. The abundance of P. major s.l. in the sampling area is further supported by the COI barcoding of females, which indicated that this species constituted 61.5% of the sandflies investigated, despite a limited number of evaluated specimens. Only one previous report on the species distribution of phlebotomines in Ankara province and the surrounding area exists (Yagci et al. 1998). In that study, the predominant species was P. perfiliewi, as identified in 98.4% of the 2248 sandflies collected from 1994–1995. However, sandfly sampling was performed in different locations (Cayirhan, Cubuk, and Kazan counties, 35–100 km north-northeast of the current sampling sites), and only one location was a peri-domestic site, where P. perfiliewi was accompanied by P. simici, P. sergenti, P. papatasi, and other species. Nevertheless, P. perfiliewi was the only phlebotomine species detected in the rest of the sampling locations, which were agricultural sites with nearby animal shelters, distant from human residential areas (Yagci et al. 1998). The frequency of P. perfiliewi and its subspecies in our study was low (4/226, 1.76%). The female predominance observed in our survey (66.1% females) was also noted in the report by Yagci's group (89.1% females). Since our field survey was confined to the neighborhoods associated with SFTV emergence, it remains to be determined whether the sandflies captured in this study represent the currently dominant sandfly species present in Ankara province.
The captured female sandflies were processed individually for the detection of phlebovirus RNA via a nested-PCR assay with degenerated primer pairs targeting the polymerase (L) segment of the viral genome (Sanchez-Seco et al. 2003). These primers are frequently used in field studies, and enables the detection of new strains, which was the case for SFTV (Carhan et al. 2010). Three sandfly specimens captured inside two cow barns (locations AI and AII) were found to possess viral RNA identified as SFTV via sequencing (Table 2). Moreover, the sequences detected in sandflies during the field survey were identical to or very similar to SFTV sequences obtained from patient samples (Fig. 2), which further supports circulation of the SFTV in the region, and symptomatic human infections caused by this strain. Interestingly, PCR-positive SFTV patients were seen in early to mid-August (data not shown), while SFTV in vectors could be identified only during early July, and samples from the same locations were negative through middle to late July. The implications of this observation, whether coincidental or not, are not clear. For a comprehensive analysis of virus circulation dynamics in the region, SFTV surveillance needs to performed throughout the sandfly season to reveal periods of peak activity, which was not possible in our study.
The phlebovirus infection rate in phlebotomines observed in this study (3/441, 0.68%) is comparable with previous reports (Moureau et al. 2010; Zhioua et al. 2010). Two of the three SFTV-infected sandflies were characterized as P. major s.l. via COI barcoding, while another phlebotomus with positive PCR could not be characterized via this method, presumably due to the presence of inhibitors, and could only be identified to the genus level (Table 2). Nevertheless, completely-characterized samples and the overall species distribution of sandflies in the field strongly suggest P.major s.l. as the vector species responsible for SFTV transmission. Another SFSV variant, Corfou virus, which is distantly related to SFTV (Fig. 2), has been detected in P. major s.l. sandflies as well (Rodhain et al. 1985). In addition to P. papatasi and P. major s.l., phleboviruses belonging to the Sicilian serocomplex have also been identified in P. ariasi (Izri et al. 2008), P. perniciosus, P. longicuspis, and Sergentomyia minuta (Zhioua et al. 2010), demonstrating the adaptation capability and wide distribution of these agents.
The evaluation of SFTV-infected sandflies individually via multiplexed PCR targeting the mitochondrial cytochrome b gene from various vertebrates enabled direct identification of the sources of blood meals. All virus-positive samples were found to be reactive for blood of bovine origin, which is not surprising since all were captured inside cow barns with livestock nearby. Two samples originating from location AII had blood meals from humans as well (Table 2), establishing another link between the virus, the vector, and human exposure in the region. The potential animal reservoirs for phleboviruses have not been studied in detail. Although preliminary findings suggested some species as candidates, the role of animals in virus survival and propagation is thought to be of lesser importance, due to the effective transovarial and venereal transmission of the virus in vectors (Tesh and Modi 1987; Tesh et al. 1992). In a recent report on TOSV exposure in animals, seroprevalence rates of 48.3% in dogs, 17.7% in goats, 32.3% in sheep, 59.6% in cats, 17.9% in cows, 22% in pigs, and 64.3% in horses were described (Navarro-Mari et al. 2011). However, virus replication could only be detected in goat serum via PCR, suggesting that these animals are amplifying hosts for the virus rather than reservoirs. Further studies are required to explore cows and other animals as possible amplifying hosts for SFTV or other viruses.
In conclusion, P. major s.l. has been identified as the vector species for the recently-discovered SFSV variant SFTV, which is associated with febrile disease with elevated liver enzymes and gastrointestinal symptoms in humans.
Footnotes
Acknowledgments
The authors are grateful to Irfan Atmaca and Salim Calis for technical assistance, and N. Emin Guven for graphics.
Author Disclosure Statement
No competing financial interests exist.