
Abstract
We detected Chikungunya virus (CHIKV) infection among wild populations of Aedes albopictus female specimens during the CHIKV outbreaks of 2009 and 2006 collected in different localities in Kerala State, India. The envelope 1 gene (E1) sequences of the virus isolate 2009 from the mosquito species showed close genetic relatedness (Kimura 2 Parameter genetic distance=0.0013) to CHIKV-positive isolates from human serum samples from the same area. E1 gene sequences from Ae. albopictus, as well as from human isolates, had the crucial non-synonymous C/T mutation at position 10670, leading to the A226V amino acid change. This natural inclination indicated the role of this mosquito species in the transmission of CHIKV during its recent outbreaks in Kerala State.

Map of Kerala State, India, showing the collection villages of mosquito specimens, including Aedes albopictus, in the study. The years of CHIKV outbreaks in the region are indicated in the map (CHIKV, Chikungunya virus).
Ae. albopictus has been implicated as an important vector for CHIKV transmission in African and European countries (Bonilauri et al. 2008; Delatte et al. 2008; Pages et al. 2009; Paupy et al. 2011). In the absence of natural detection of CHIKV in this species from India, thus far its role in transmission was indirectly related to its abundance, and human–vector contact (Kumar et al. 2008; Eapen et al. 2010).
The A226V mutation in the E1 gene of the virus has been shown to be advantageous for the virus to undergo development and dissemination in the vector species Ae. albopictus (Tsetsarkin et al. 2007). The virus strains isolated from human blood samples during the 2006 outbreaks in Kerala and Karnataka states did not have this mutation (Kumar et al. 2007; Arankalle et al. 2007), and the worst affected regions in 2006 were the eastern districts of Karnataka, and the coastal regions in Kerala (Alappuzha District), where Ae. albopictus is less abundant. However, since 2007, all of the isolates of CHIKV reported from Kerala and Karnataka States were found to have this mutation (Kumar et al. 2008; Santhosh et al. 2009). This advantageous mutation (Tsetsarkin et al. 2007) may have been repaired by natural selection in the virus population on the western side of Western Ghats in Kerala and Karnataka States, which harbor an abundant population of Ae. albopictus (Kumar et al., 2011). This mid-highland semi-forested region is cultivated with crops such as rubber, pineapple, and areca, and this setting favors the proliferation of Ae. albopictus (Kumar et al. 2008; Eapen et al. 2010). The micro-evolutionary adaptation of CHIKV may have played a major role in the widespread outbreak of Chikungunya fever in the region, contributing about 59.4% of the total cases seen in India from 2006–2011. Isolates from other regions of the country did not record the A226V mutation (Sudeep et al. 2011; Grandadam et al. 2011).
As a part of our CHIKV epidemic investigations, extensive entomological surveys were carried out in affected areas in Kasaragod District of Kerala and Dakshin Kannada District of Karnataka during 2008, and in Kozhikode District, Kerala, during 2009. Also, vector surveillance is being carried out on a continuing basis in sentinel and randomly-selected villages in the central region of Kerala (the districts of Alappuzha, Kottayam, Pathanamthitta, and Idukki), where major outbreaks of CHIKV occurred during 2006 and 2007 (Fig. 1). We processed these Ae. albopictus and other mosquito specimens for CHIKV infection as follows. The mosquitoes were transported alive in 2×2-foot Barraud cages from the collection sites to the laboratory. They were identified and female specimens were categorized into unfed (UF), fully-fed (FF), semi-gravid (SG), or gravid (G), based on abdominal condition. Specimens with the same abdominal condition collected from a given area were pooled together and processed by reverse transcriptase polymerase chain reaction (PCR) assay to detect viral infection. The pool size varied from 1 to 10 specimens, the maximum pool size being 5 for FF, SG, and G females, and 10 for UF females and male specimens. Total RNA was extracted using TRI reagent RT (Molecular Research Center, Cincinnati, OH). Reverse transcriptase PCR was performed using a Titan 1-tube RT-PCR kit (Roche Diagnostics, Mannheim, Germany), using CHIKV-specific non-structural protein 1 (nsP1) DNA primers (Hasebe et al. 2002).
A total of 1204 field-collected female specimens of Ae. albopictus from 33 villages were processed in 400 pools for CHIKV infection from 2006–2010 (Table 1). Among these, one pool of 5 SG Ae. albopictus from Valayam village, Kozhikode (11.752450° N, 75.701250° E), collected during the 2009 CHIKV outbreak, and another one with 3 FF Ae. albopictus specimens collected from Kadakkarappalli village (9.686499° N, 76.321142° E), Alappuzha, Kerala State, during the 2006 outbreak, were found to be positive for CHIKV infection. The envelope 1 gene (E1) of CHIKV from the positive pool of SG specimens was PCR amplified (Kumar et al. 2008) to check for the A226V mutation. The sequence was deposited with the GenBank (accession no. HQ599560). The specimens of this pool were collected outdoors from a rubber plantation in the village. Serum samples were also collected during the survey from the same region during the 2009 outbreak. Among these, 22 CHIKV-positive samples were recorded. The infection recorded in FF specimens collected from Alappuzha District in Kerala may not provide a definitive vectorial role for Ae. albopictus. Hence only the diagnostic nsP1 gene amplified was sequenced to confirm the infection status (GenBank accession no. JQ013788). Parallel human serum samples collected from this region during the 2006 outbreak did not have the A226V mutation in the E1 gene (Kumar et al. 2007).
One pool of FF specimens collected from Kadakkarappalli, Alappuzha District, and banother pool of SG specimens from Valayam, Kozhikode District, were found to be positive for CHIKV infection.
UF, unfed; FF, fully-fed; SG, semi-gravid; G, gravid; CHIKV, Chikungunya virus.
None of the other species of mosquito specimens processed (Mansonia uniformis [64], Culex vishnui gp. [18], Armigeres subalbatus [13], Cx. tritaeniorhynchus [12], and Ae. aegypti [6]) were found to be positive for CHIKV infection.
The E1 gene sequences of CHIKV isolated from Ae. albopictus in this study, and the viral isolates from human samples from India (2006–2010), available at the GenBank were subjected to maximum likelihood (ML) phylogenetic analysis using MEGA 5.0 (Tamura et al. 2010), incorporating representatives of ECSA and ASIAN genotypes of the virus (Fig. 2). Three CHIKV parallel-positive serum samples collected from the same locality (Valayam) were also included in this analysis. The ML phylogenetic consensus tree deduced with bootstrapping of 500 replications indicated that the CHIKV isolate from Ae. albopictus belonged to the same lineage as the human sample isolates collected during the 2009 Kozhikode outbreak (Kimura 2 Parameter genetic distance=0.0013), and had the amino acid valine in the 226 position of the E1 gene.
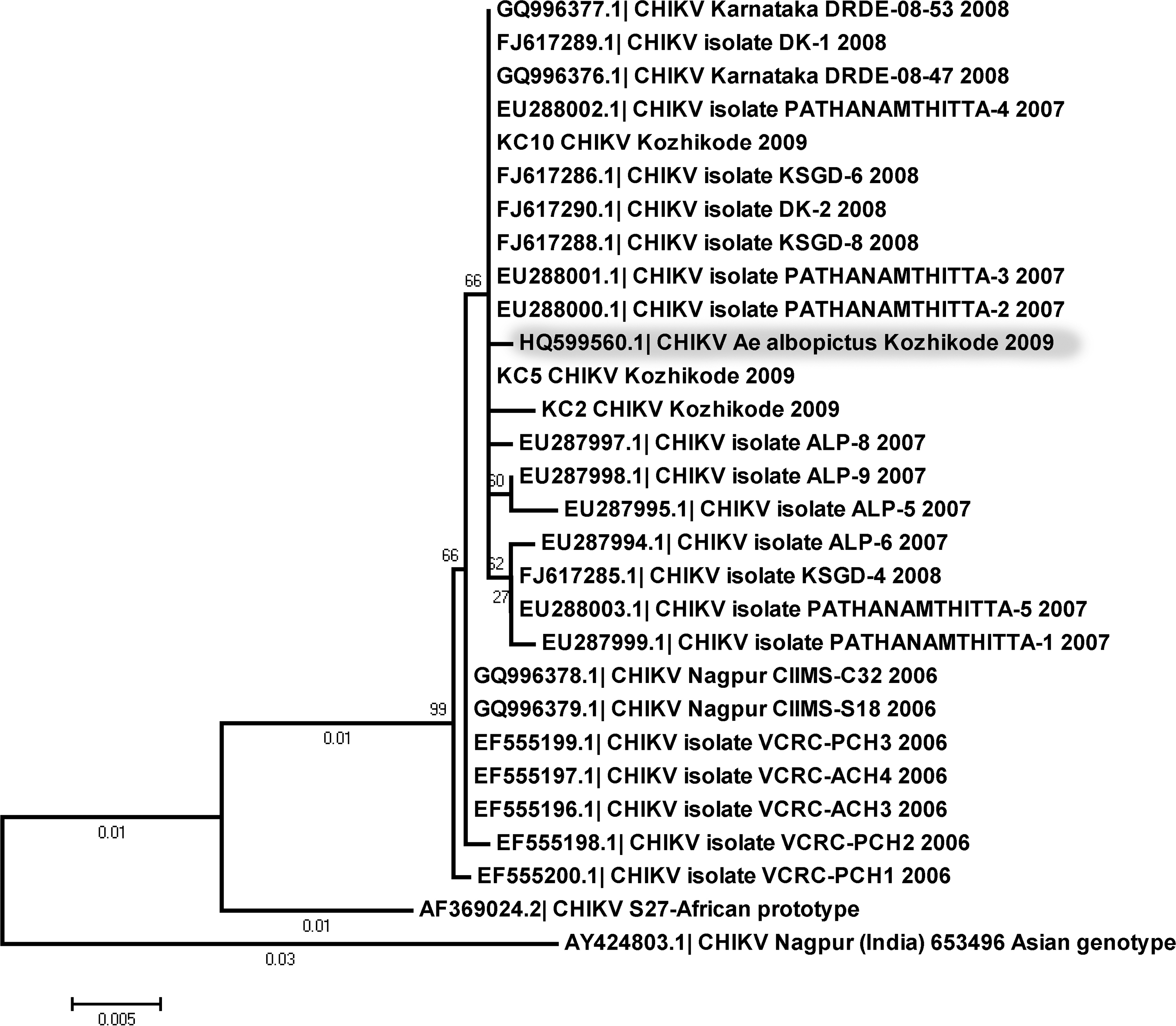
Phylogenetic relationship of Chikungunya virus (CHIKV) isolates based on maximum likelihood analysis of the envelope 1 gene sequences. Bootstrap values for 500 replications are provided in the nodes. The CHIKV isolate from Aedes albopictus is shaded grey. KC-2, KC-5, and KC-10 are CHIKV-positive human samples included in the analysis that were collected from the Kozhikode region during the outbreak of 2009. Their GenBank accession numbers are JQ886092, JQ886093, and JQ886094, respectively.
None of the 658 female specimens of Ae. albopictus processed from sentinel villages (where an epidemic outbreak of Chikungunya fever occurred during 2006–2007), during the post-epidemic surveillance period (2008–2010) yielded positive reactions to CHIKV, indicating a lack of its circulation in the area during this period. Also, 143 male Ae. albopictus specimens that emerged from immature collections, in both epidemic outbreaks and post-epidemic situations, were tested and none was found to be positive for CHIKV infection. The absence of virus in this field-collected male population is an indicator of a lack of vertical (trans-ovarial) transmission of CHIKV in Ae. albopictus. Similar observations of the absence of trans-ovarial transmission of CHIKV were recorded in India and Indian Ocean islands (Mourya 1987; Vazeille et al, 2009). A recent study in Italy (Bellini et al, 2012), which showed small numbers of cases of CHIKV vertical transmission among the Ae. albopictus specimens analyzed (0.43%), concluded that vertical transmission of CHIKV is a rare event, a finding in agreement with another unpublished field study carried out there, in which researchers analyzed about 8000 immature specimens and none were found to be positive for CHIKV infection.
CHIKV infection in SG specimens of Ae. albopictus, and its genetic relatedness with virus isolated from human samples from the same area, indicated a role of Ae. albopictus in the transmission of CHIKV during the recent outbreaks in the Kerala and Karnataka States of India. Vertical transmission could not be demonstrated, as none of the freshly emerged UF female and male specimens of this species had CHIKV infection during the post-epidemic period. Also, male specimens during epidemic outbreaks did not appear to have CHIKV infection.
Footnotes
Acknowledgments
The authors are grateful to Mr. P.M. Ajithlal, Mr. N. Krishnamoorthy, Mrs. Jessu Mathew, and Ms. Kochurani George, for technical assistance rendered for this study.
Author Disclosure Statement
No competing financial interests exist.