
Abstract
Human lymphatic filariasis is caused by filarial worms such as Brugia malayi for which the major reservoir is domestic cats. However, domestic cats or dogs also carry nonhuman filaria such as Brugia pahangi and Dirofilaria immitis. We have developed a single-tube, real-time PCR with a high-resolution melting (HRM) analysis assay for detection and identification of B. malayi, B. pahangi, and D. immitis in blood samples. The designated primer pair in the PCR can amplify a 114-bp region of mitochondrial 12S rRNA genes of these filarial worms. Subsequently, the HRM assay showed a specific melting temperature for each species. The assay showed the highest sensitivity and specificity in comparison with DNA sequences after assessment with 34 cat and 14 dog blood samples. This assay could be helpful for epidemiological studies of reservoirs and vectors.
Introduction
Brugian filariasis, a human filarial infection caused by B. malayi, has been targeted by the World Health Organization (WHO) to be eliminated by the year 2020 (Ottesen 2002). Assessing the burden of filarial parasites is one of the important strategies for the control of filariasis. However, control programs in endemic areas of subperiodic B. malayi have had difficulty in eliminating infection due to continuing zoonotic transmission from reservoir hosts (Lim and Mak 1983).
In Thailand, brugian filariasis is endemic in the south, where domestic cats serve as the major reservoir (Filariasis Division 2009). Domestic cats carry not only B. malayi but also B. pahangi and D. immitis. Identification of filarial species is done conventionally using microfilaria morphology (Yen and Mak 1978, Chansiri et al. 2002). The application of molecular technology could have a great impact when it is necessary to distinguish closely related species or subspecies. Seminested PCR–restriction fragment length polymorphism (PCR-RFLP) analysis of the internal transcribed spacer 1 (ITS1) has been used to differentiate among B. malayi, B. pahangi, and D. immitis (Nuchprayoon et al. 2003). However, these methods are relatively tedious and complicated.
High-resolution melting–real-time PCR (HRM-PCR), introduced by Wittwer (Wittwer et al. 2003), is one of the fastest real-time PCR systems. The advantages of the HRM method are that amplification and detection steps can be conducted in the same reaction tube or well and no post-PCR processing is required (Dobrowolski et al. 2009, Nettuwakul et al. 2010). The master mix used in HRM contains a DNA-binding dye that stains target sequences and results in sharp melting signals that are analyzed using the Gene Scanning Software in the HRM instrument. (Dobrowolski et al. 2009).
In the present study, we designed a HRM-PCR assay targeting the mitochondrial partial 12S rRNA gene, which is highly conserved and contains genus- and species-specific sequence variations useful for identifying the 3 filaria species, i.e., B. malayi, B. pahangi, and D. immitis, in blood samples from cats and dogs.
Materials and Methods
Reference strains and blood samples
Reference strains for the HRM assay development were B. malayi, B. pahangi, and D. immitis, which were provided from TRS Labs (Athens, Georgia). Thirty-four blood samples of cats from endemic areas of brugian filariasis in Narathiwat and Surat Thani provinces as well as 14 dog blood samples from the Bangkok area were collected. Three thick blood smears on glass slides were prepared from 2–3 drops of blood for each sample and kept at room temperature after air drying. Plasmodium falciparum–positive human blood samples and 2 Babesia spp.–positive dog blood samples were also recruited for specificity testing of the assay.
Microfilaria detection in the thick blood smear preparation
The thick blood smears on slides were dehemoglobinized by briefly dipping the slide in water for hemolysis. One slide of a sample was examined for microfilariae under a microscope using Giemsa staining (World Health Organization 1991).
DNA extraction
The blood film was released from the glass slide by dropping 100 μL of Tris-EDTA buffer onto the dehemoglobinized thick blood smear and incubating it for 5 min at room temperature. The solution was transferred to a 1.5-mL microcentrifuge tube and centrifuged at 12,000 rpm for 10 min. The upper solution was discarded, and the pellet was suspended in 200 μL of phosphate-buffered saline (PBS) for DNA extraction using the High Pure® PCR Template Preparation Kit (Roche Applied Science, Germany) according to the manufacturer's instructions. DNA was eluted in 100 μL of elution buffer and stored at −20°C until use. The DNA concentration was measured using a spectrophotometer (Nanodrop 1000, Thermo Scientific).
Primers
The primer designs were based on alignments of the mitochondrial partial 12S rRNA genes of B. malayi, B. pahangi, and D. immitis; these genes contain genus- and species-specific sequence variations useful for identifying the 3 filaria species (Fig. 1). Sequences were obtained from the National Center for Biotechnology Information database (

Alignment of partial mitochondrial 12S rRNA gene sequences of Brugia pahangi (GenBank accession number AM779851; positions 154–264), B. malayi (GenBank accession number AJ544843; positions 177–287), and Dirofilaria immitis (GenBank accession number FN391554, positions 161–275). Dots indicates identity; dashes indicate deletion from the above consensus sequence; and gray areas indicate primers.
HRM real-time PCR assay
PCR and HRM were performed in a single run on a LightCycler LC480 instrument (Roche Diagnostics, Penzberg, Germany) in a reaction mixture containing 3 μL of DNA, 0.25 pmol of each primer, and 3 mM MgCl2 in 5 μL of the LightCycler 480 High-Resolution Melting Master mixture containing ResoLight dye (Roche Diagnostics) with PCR-grade water adjusted to a final volume of 10 μL. Nuclease-free water replaced the DNA template for negative controls. Positive controls were prepared by adding 3 μL of template DNA from reference species of B. malayi, B. pahangi, and D. immitis.
Reaction conditions included an activation step at 95°C for 5 min followed by a 40-step amplification of 10 s at 95°C, 10 s at 58°C, and 10 s at 72°C. Subsequently, the products were heated to 95.8°C for 1 min and then cooled to 40°C for 1 min. HRM was performed from 65°C to 95°C, rising at 1°C/s with 25 acquisitions per degree. The final cooling step was 40°C for 10 s.
Melting curves were generated, normalized, temperature-shifted, and converted to difference plots by LightCycler® 480 gene scanning software (Roche Applied Science, Germany). Also, various concentrations of genomic DNA from the same samples of the 3 species were amplified and used to determine % coefficient of variation (%CV) of the melting temperatures (Tms).
Determination of sensitivity and specificity
The lowest levels of detection of B. malayi and D. immitis were determined in 2-fold serial dilutions of the genomic DNA. To verify the reproducibility of the assay, DNA from each reference positive control was tested on 10 different days. Mean, standard deviation (SD), and %CV were calculated.
DNA sequencing
The PCR products of the samples were further purified using a High Pure PCR Product Purification kit (Roche, Germany). They were submitted for DNA sequencing. The sequence alignment and sequence analysis were performed with Clustal W (Thompson et al. 1994).
Animal ethics
The study protocol was approved by the Animal Ethics Committee of Faculty of Medicine Siriraj Hospital, Mahidol University, based on the Ethics of Animal Experimentation of the National Research Council of Thailand (Reference No. SI-ACUP 009/2552).
Results
The reference strains of B. malayi, B. pahangi, and D. immitis could be recognized by the HRM assay at 10 times the reproducibility of Tms at 75±0.25, 77.46±0.24, and 74.13±0.28°C, respectively, with %CV at 0.31%, 0.33%, and 0.38%, respectively. Figure 2 presents melting curve analyses as well as the normalized and temperature-shifted difference plots.

Melting peaks (
The assay could detect as little as 19.4 pg/reaction and 16.4 pg/reaction of B. malayi and D. immitis genomic DNA, respectively. The results were negative when tested with P. falciparum and Babesia spp. blood samples.
HRM assay of blood samples
By using the HRM assay as a tool to identify microfilaria in 48 blood samples, 35 samples were positive. They were 18 B. pahangi (Tm at 77.30°C to 77.89°C), 8 B. malayi (Tm at 75.71°C to 76.28°C), and 9 D. immitis (Tm at 73.85°C to 74.64°C). The results are presented in Fig. 3 and summarized in Table 1. DNA sequences were analyzed in 14 of the 35 HRM-positive samples and the 3 references species. All results observed were concordant.
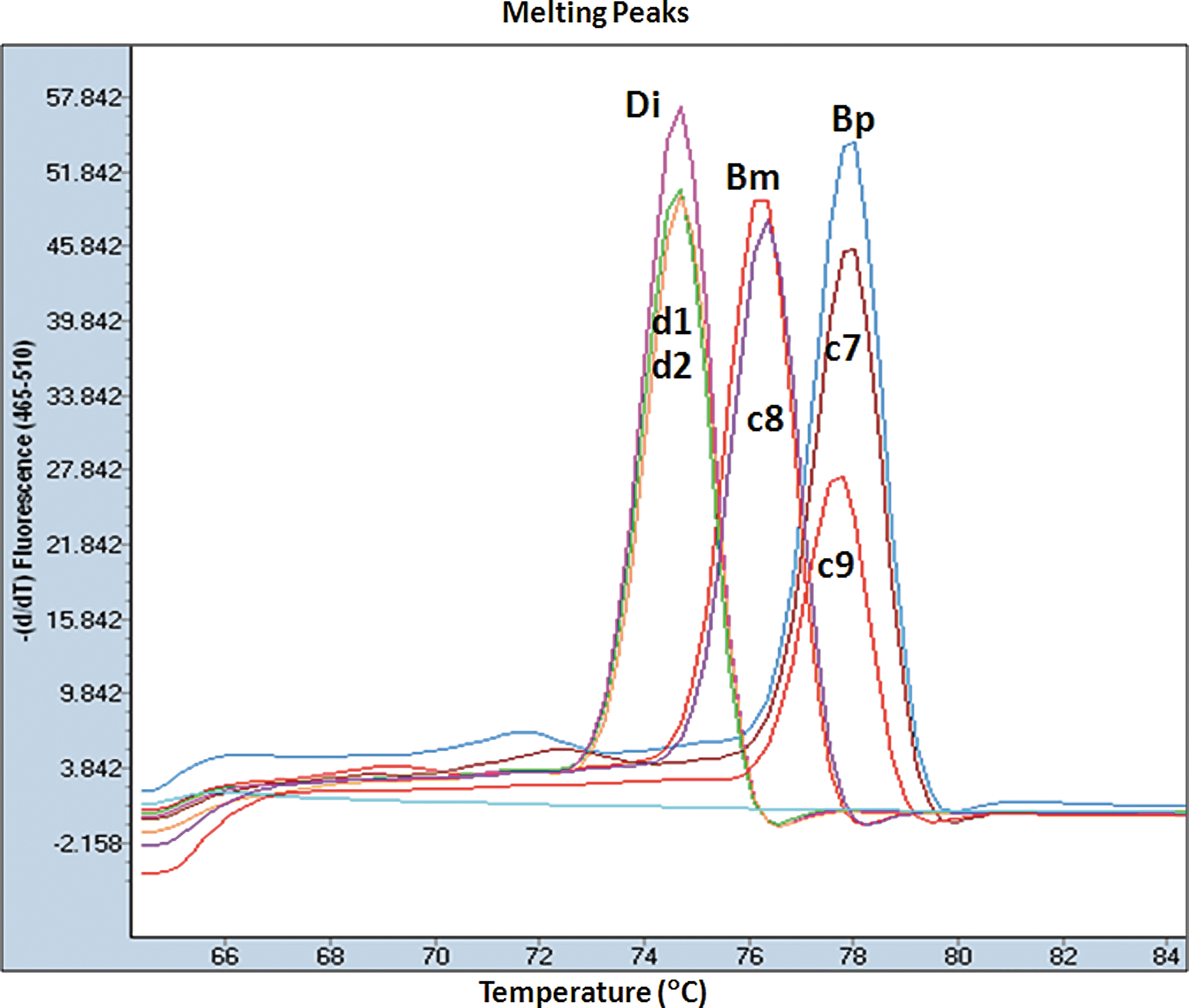
Melting peaks of the high-resolution melting (HRM) results of the dog samples (d1 and d2), cat samples (c7, c8, and c9), and control species (Bm, Brugia malayi; Bp, B. pahangi; and Di, Dirofilaria immitis). Color images available online at
Tm, Melting temperature.
Discussion
To control a filarial disease effectively, assessment of the nature and scope of the parasite problem in an endemic area is needed; however, collection of detailed and accurate epidemiological data is hampered by difficulties in detecting and identifying filarial parasites in human, animal, and vector populations (Denham and McGreenvy 1977).
We designed the HRM assay for detection and differentiation of the 3 filaria species, i.e., B, malayi, B. pahangi, and D. immitis, in blood samples of cats and dogs. The HRM assay detected the partial mitochondrial 12S rRNA gene of these species by using a single primer pair that amplified 144 bp of the gene. The assay can identify the PCR product by differentiation of the melting peak base on 3–5 different bases in the amplicon. Each double-stranded DNA (dsDNA) fragment has its characteristic melting behavior, which depends mainly on its GC content, length, and sequence composition (Ririe et al. 1997).
In our study, Tm analysis was essential in identifying the three filaria species. The normalized and temperature-shifted difference plots also divided the species into 3 groups. Thus, the 3 filaria species could be clearly distinguished by both using the Tm as well as the normalized and temperature-shifted difference plot. Moreover, the developed HRM could detect as little as 1 microfilaria/reaction or a microfilaria/60-μL blood sample. The sensitivity of the developed assay could be increased if a greater amount of blood is used, for example, by extracting DNA from 2–3 thick blood smear slides in one reaction.
The assay also shows high specificity. No false-positive results were detected in either P. falciparum, Babesia spp., or parasitic-free cat or dog samples. Moreover, DNA sequencing of the study samples shows them to be 100% identical with the HRM result, thus confirming that HRM is as effective as gene sequencing. Vossen et al. stated in their study that HRM analysis is more effective than full gene sequencing, even for genes with small amounts of exons (Vossen et al. 2009). The %CV of Tms of the three reference species, confirming the reproducibility of the test, was very high.
A real-time fluorescence resonance energy transfer (FRET) PCR combined with melting curve analysis has been developed for differentiating B. malayi and B. pahangi DNA in blood samples. However, our HRM assay uses one set of primers and fluorophore-labeled hybridization probes specific for HhaI repetitive DNA (Thanchomnang et al. 2010). The HRM reaction can be performed in a single tube (or well) using a single pair of primers, and no specific probes are needed. The assay can be run rapidly with large sample sets, such as 96 or 384 samples, in one run of a PCR, using approximately 5 min of melting and analysis. The cost effectiveness of HRM is comparable to or cheaper than conventional PCR and sequencing (Dobrowolski et al. 2009, Zhou et al. 2005). In addition, our method of DNA extraction from a dry thick blood smear was simple.
In the samples studied here, we did not find more than 1 Tm of mix infection. In this case, experiments of mix DNA from samples were undertaken, as shown in Table 2. The assay could not demonstrate all 2 or 3 peaks of the mix species from the effect of PCR competition. However, Tm values of the heteruduplex amplicon were present at 67.33° to 68.5°C in the mix D. immitis and Brugia spp. and at 69.14°C to 71.81°C in the mix B. malayi and B. pahangi (Fig. 4).
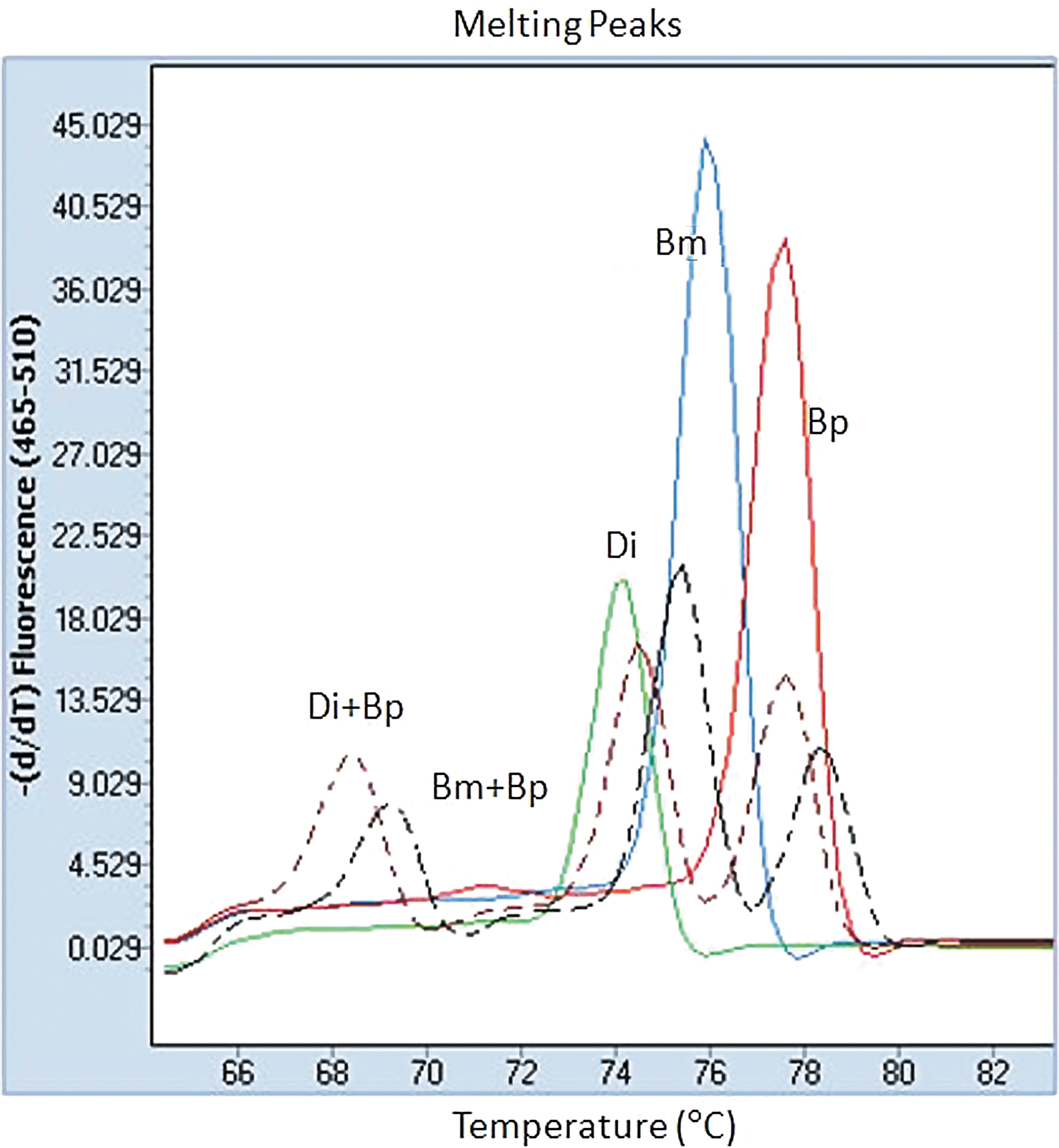
Melting peaks of high-resolution melting (HRM) results of mix Brugia malayi and B. pahangi DNA at 10:1 (black dotted line) and mix Dirofilaria immitis and B. pahangi DNA at 1:10 (brown dotted line). Two separate fluorescence peaks at different melting temperatures of species and heteruduplex melting peaks are present at 68.5°C and 69.14°C. (Bm, B. malayi; Bp, B. pahangi; and Di, D. immitis). Color images available online at
Tm, Melting temperature; Bm, B. malayi; Bp, B. pahangi; Di, D. immitis.
Conclusion
The HRM assay is a reliable and cost-effective method for rapid identification of B. malayi, B. pahangi, and D. immitis. It will also be useful for many other applications, including adaptation for field-based technology, such as epidemiological surveys in reservoir hosts and vectors.
Footnotes
Acknowledgments
This study was supported by grants from Faculty of Medicine Siriraj Hospital, Mahidol University. Thanks are also extended to Mr. Choochai Nettuwakul, Mr. Wittaya Klongkaew, and Mr. Supit Yodmek for their assistance. We thank you Mr. William M Honsa, Jr., for grammatical proofing of this manuscript.
Author Disclosure Statement
No competing financial interests exist.