
Abstract
Here we report a case of co-infection with Orientia tsutsugamushi, the causative agent of scrub typhus, and Arsenophonus nasoniae in a woman with a rash and an eschar who returned from a trip to Southeast Asia. A. nasoniae was previously considered to be a secondary insect and tick endosymbiont of unknown pathogenicity in humans. We amplified both O. tsutsugamushi and A. nasoniae DNA from a skin eschar with qPCR, and a seroconversion for O. tsutsugamushi and A. nasoniae was observed with immunofluorescence assays and western blotting for this patient. And we used 2-D western blotting with an A. nasoniae antigen and polyclonal mouse anti-A. nasoniae antibodies produced in our laboratory to detect the specific antigenic A. nasoniae proteins.
Introduction
Orientia tsutsugamushi is an obligate intracellular bacterium belonging to the α-subdivision of proteobacteria, family Rickettsiaceae, and is responsible for scrub typhus. Scrub typhus is endemic across extensive parts of southern and southeastern Asia, Australia, and the Pacific region. O. tsutsugamushi is transmitted to humans by the bites of larval trombiculid mites (chiggers). The larvae typically bite humans on the lower extremities or in the genital region. The bacterium remains a major cause of undifferentiated fever in Asia. The severity of the disease varies from asymptomatic to fatal. With the growing popularity of travel in Asia, more travelers are returning to nonendemic areas with this disease. Co-infection with scrub typhus and others pathogens such as dengue virus, Japanese encephalitis virus, and Leptospira has been detected in Southeast Asia (Wang et al. 2003, Suttinont et al. 2006, Lee and Liu 2007).
We tested skin biopsies received by our national reference center (URMITE, Marseille, France) to diagnose arthropod-borne bacterial diseases by PCR, including potential emerging bacteria as A. nasoniae. We detected a case of co-infection with A. nasoniae and O. tsutsugamushi in a French patient who had recently traveled to Southeast Asia. Serological testing confirmed co-infection in the patient, but the role that A. nasoniae may have played in disease pathogenesis is still unknown.
Material and Methods
Sample collection
As a National Reference Center for Rickettsioses, Bartonelloses, and Anaplasmoses, we routinely receive blood, skin biopsy, and serum specimens from patients bitten by ticks or others arthropods. We have been routinely and systematically screening 67 skin biopsy samples for the presence of A. nasoniae DNA in 2010, then 88 in 2011. The specific PCR for O. tsutsugamushi was performed only on skin biopsy from a patient who had recently traveled in Southeast Asia; 7 patients were tested during the year 2010 and 2011.
PCR assay
DNA was extracted from patient samples using a QIAamp tissue kit and a QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany) for eschar biopsy and blood samples, respectively. Each eschar biopsy sample was screened by qPCR for O. tsutsugamushi using primers and probes targeting a 47-kD outer membrane protein gene as previously described (Jiang et al. 2004). The reaction was conducted in a Light Cycler instrument (Roche Diagnostics GmbH, Germany). Primers and probes targeting a specific region of the ftsY and rpoB genes of A. nasoniae were used for DNA-based detection of A. nasoniae (Table 1) (Mediannikov et al. 2012). For each test, two negative controls (sterile water and DNA from a sterile skin biopsy specimen) and a positive control (DNA extracted from A. nasoniae strain IZ40 cultured in blood agar) were included.
Serology
Serum specimens were tested via microimmunofluorescence assays using a large panel of antigens, including O. tsutsugamushi antigens (Gilliam, Karp, Kato, and Kawasaki strains), as previously described (La Scola and Raoult 1997). Titers of ≥1:128 for immunoglobulin G (IgG) and ≥1:64 for IgM and/or seroconversion were considered evidence of recent infections.
A. nasoniae antigen was produced in our laboratory using the IZ40 A. nasoniae strain, isolated from Ixodes ricinus (Mediannikov et al. 2012). This strain was grown on blood agar at 28°C in a 5% CO2 incubator. After incubation for 5 days, the bacteria were harvested and suspended in sterile distilled water prior to being frozen at −20°C. All microimmunofluorescence assays were performed with the same protocol. Polyclonal A. nasoniae antibodies were produced in 6- to 8-week-old female BALB/c mice that were inoculated intraperitoneally with 106 colony-forming units (CFU)/mL of A. nasoniae suspended in CpG adjuvant. The sera from these infected mice were used as positive controls for serology.
Western blotting
Western blotting was performed as previously reported (La Scola and Raoult 1997) using A. nasoniae (IZ40 strain) and O. tsutsugamushi (Gilliam strain) antigen as described above and the four serum samples collected from the patient. We used as positive control A. nasoniae (IZ40 strain) and O. tsutsugamushi (Gilliam strain) polyclonal mouse antibodies produced in our laboratory.
2-D electrophoresis
Sample preparation for 2-D electrophoresis
A. nasoniae was cultured on blood agar for 3 days at 28°C in a 5% CO2 incubator before being harvested and ultracentrifuged (100,000×g for 2 h at 4°C in a Beckman MLS-50 rotor). The bacterial proteins were precipitated using a PlusOne 2-D Clean-Up Kit (GE Healthcare, Chalfont St. Giles, UK). The resulting pellet was resuspended in solubilizing buffer. The protein extract was quantified using a Bio-Rad DC assay.
2-D electrophoresis and silver staining
Immunoblotting was performed on 13-cm strips, and spots on 18-cm strips were submitted for mass spectrometry analysis. Immobiline™ DryStrips, pH 3–10 (GE Healthcare, Chalfont St. Giles, UK) were rehydrated overnight with 100 μg of protein/13-cm strip or 150 μg of protein/18-cm strip in rehydration solution supplemented with 0.5% (vol/vol) IPG buffer (pH 3–10) (GE Healthcare, Chalfont St. Giles, UK). Isoelectric focusing (IEF) was conducted according to the manufacturer's instructions (Ettan IPGphor II, GE Healthcare, Chalfont St. Giles, UK). Prior to gel electrophoresis, the strips were equilibrated as previously described (Renesto et al. 2005). The strips were then embedded in 0.5% agarose, and the proteins were resolved with a 10% acrylamide gel (Ettan™ DALT, GE Healthcare, Chalfont St. Giles, UK) for the 18-cm strips and a 9–16% acrylamide gel for the 13-cm strips (Bio-Rad Protean Mini xi chamber), using the manufacturer's suggested protocol. For protein identification following electrophoresis, the gels were silver-stained (Nesterenko et al. 1994) and digitized by transmission scanning (ImageScanner, GE Healthcare, Chalfont St. Giles, UK).
Western blot assays
The proteins resolved on the 2-D gels were transferred onto a nitrocellulose membrane in a transblot cell (Bio-Rad) for 1.5 h. The membranes were then blocked with Tris-buffered saline with Triton X-100 (TBST; 50 mM Tris-HCl [Invitrogen Cergy-Pontoise, France] and 250 mM NaCl supplemented with 0.5% Triton X-100) and 5% nonfat dried milk overnight. Then, the membranes were incubated with the polyclonal mouse anti-A. nasoniae antibodies in a solution of TBST and 5% nonfat dried milk (dilution of 1:500). Following a 1-h incubation, the membranes were washed three times with TBST and incubated with peroxidase-conjugated immunoglobulin (goat anti-mice IgG [H+L] at a 1:1000 dilution; Southern Biotechnology, Birmingham, AL). Each membrane was treated with the secondary antibody and washed three times, as previously described. Detection was performed using a commercially available chemiluminescence kit (ECL™ Western Blotting Analysis System, GE Healthcare). The membranes were exposed to Hyperfilm™ ECL (GE Healthcare, Chalfont St. Giles, UK) and subsequently developed using an automated film processor (Hyperprocessor™, GE Healthcare, Chalfont St. Giles, UK).
In-gel digestion and matrix-assisted laser desorption/ionization–time-of-flight mass spectrometry
The protein spots were manually excised from the silver-stained gels, destained, and subjected to in-gel digestion with trypsin (Shevchenko et al. 1996). The tryptic peptides were extracted from the gel via a successive treatment with 80% acetonitrile in 0.2 % trifluoroacetic acid (TFA). The extracts were dried at ambient room temperature. The peptides were co-crystallized in the presence of 0.5% TFA onto the matrix-assisted laser desorption/ionization (MALDI) target plate with an equal amount of matrix solution (3 mg/mL−1) consisting of alpha cyano-4-hydroxycinnamic acid and 50% acetonitrile. Mass spectrometric analyses were performed using a MALDI-time of flight (TOF)/TOF Bruker Ultraflex II spectrometer (Bruker Daltonics, France). The spectra were calibrated internally using trypsin autolytic peptides. To identify the proteins, the peptide mass lists were matched against databases proposed by the Mascot search engine (Matrix Science TD, UK; available at
Results
Case report
In December, 2010, a previously healthy 31-year-old woman was admitted to the infectious diseases unit of a hospital in Vannes, France, presenting with oscillating fever, shivers, and myalgia associated with violent and persistent headache without meningeal signs but with confusional syndrome for 1 week. Three weeks earlier, she had returned from a 2-week trip to Cambodia where she had spent some nights in the jungle. She had followed the usual recommendations to protect against malaria. She took atovaquone and proguanil 1 day before her departure until 1 week after her return. Her malaria test, a blood smear stained with Giemsa, was negative. Blood, stool, and urine cultures also tested negative for parasites. Her cardiopulmonary examination was normal. Her leukocyte count was 9.7 G/L, with 19% of her lymphocytes appearing atypical, similar to lymphocytes found in blood mononucleosis syndrome. Her platelet level was 174 G/L. She presented a biologic inflammatory syndrome with elevated C-reactive protein and elevated fibrinogen. Her liver enzyme blood levels were slightly elevated (aspartate amino transferase, 130 UI/L; alanine amino transferase, 146 UI/L; γ-glutamyl transpeptidase, 140 UI/L). Upon subsequent questioning, the patient acknowledged that she was bitten by an insect during her trip to Asia. A skin examination revealed a black maculopapular lesion in the right popliteal fossa (Fig. 1). A skin biopsy was performed and was sent to our laboratory, as were as serum and blood samples. The patient was clinically diagnosed with rickettsioses and a treatment with 200 mg/day of doxycycline was started. She rapidly improved and her symptoms disappeared within 48 h.

Inoculation eschar, a black maculopapular lesion in the right popliteal fossa. Color images available online at
Laboratory results
O. tsutsugamushi and A. nasoniae DNA were detected in the skin biopsy using primers and probes targeting a 47-kD outer membrane protein gene of O. tsutsugamushi and the ftsY and rpoB genes of A. nasoniae. PCR analysis of the patient's blood sample was negative for the two bacteria. The sequence (120 bp) of the rpoB gene obtained from the skin biopsy exhibited 98.4% homology with A. nasoniae (GenBank accession no. FN545258).
IFA was performed for both O. tsutsugamushi and A. nasoniae on two early sera taken at the onset of the symptoms and 1 week later during the hospitalization and two convalescent sera sampled 1 month after she was hospitalized and 15 days later. Seroconversion was observed in the patient for O. tsutsugamushi and A. nasoniae (Table 2). In fact, the first serology tests performed at the onset of symptoms and during the hospitalization were negative for both O. tsutsugamushi and A. nasoniae. Sera collected 1 month after hospitalization showed an IgG titer of 1:64 and IgM titer of 1:64 with O. tsutsugamushi serotype Gilliam antigen. For A. nasoniae, an IgG titers of 1:128 (Fig. 2) were observed in the fourth serum. To confirm the serological results, western blots were performed with the patient's sera using mouse anti-A. nasoniae sera as described above. The western blot confirmed A. nasoniae and O. tsutsugamushi were the causative agents (Fig. 3) on the third and fourth sera; the first and second sera were negative.

Seroconversion observed with immunofluorescence assay using O. tsutsugamushi and A. nasoniae antigens and the patient's sera. Magnification, 100×. (
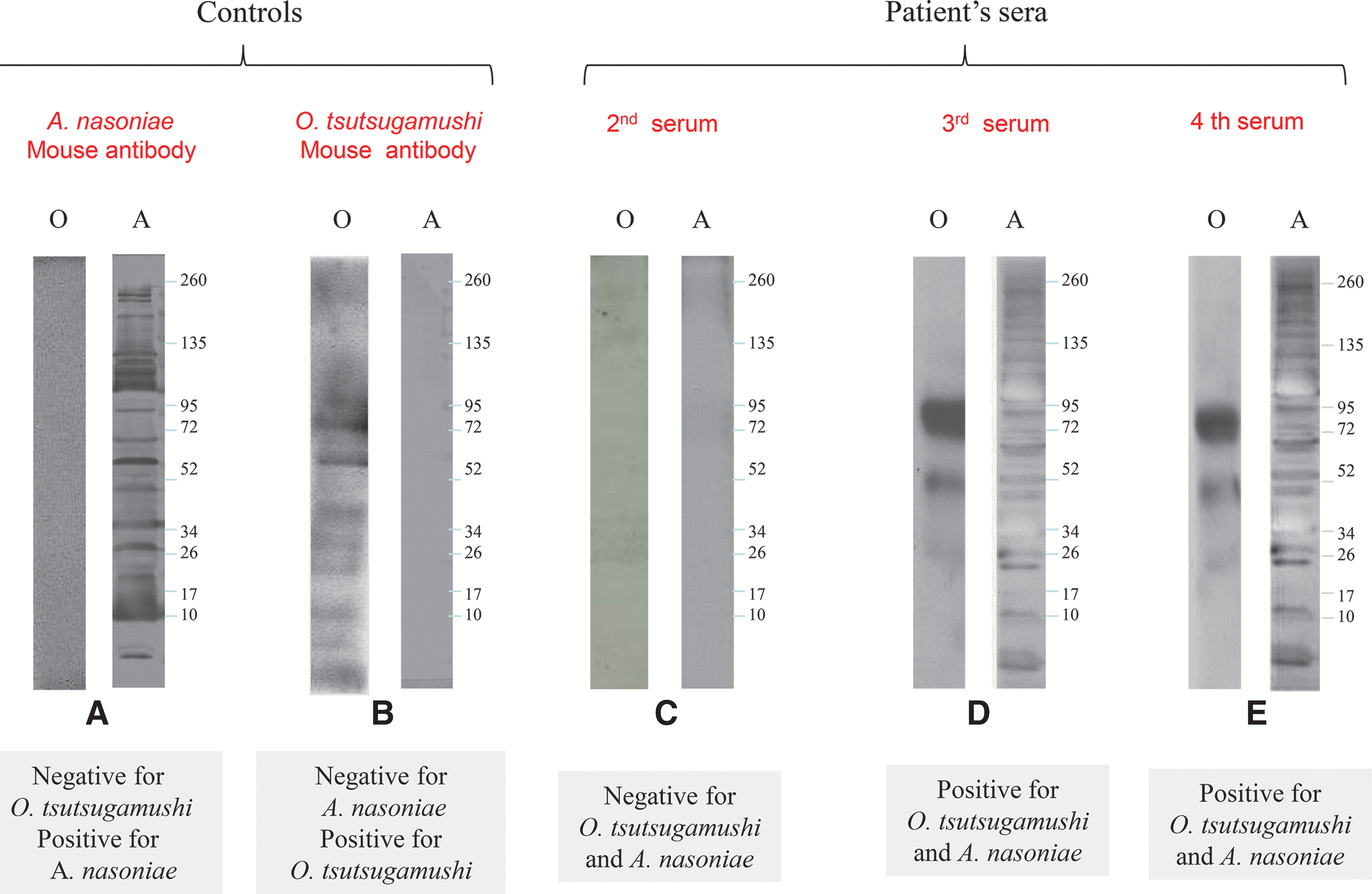
Western blot of O. tsutsugamushi (O) and A. nasoniae (A) antigen: (
IFA, immunfluorescence assay; ND, not done; IgG, immunoglobulin G; IgM, immunoglobulin M.
Identification of antigenic A. nasoniae proteins
All immunoreactive discriminated spots identified in reaction with mouse antibodies were manually excised from the gels, and each spot corresponded to a distinct protein (Fig. 4). The 2-D immunoproteomic profile of the mouse serum (Fig. 4B) was compared with the A. nasoniae proteome (Fig. 4A). The antigenic proteins were identified from the corresponding silver-stained gel. All of the proteins labeled in the gel are listed in Table 3. In total, six proteins were identified successfully. Six proteins detected by the polyclonal anti-A. nasoniae antibody were identified by MALDI-TOF analysis as OmpA, a 10-kD chaperonin, an adenylate kinase, a protein providing DNA protection during starvation, serine hydroxymethyl transferase, and a periplasmic protein disulfide isomerase (Table 3).
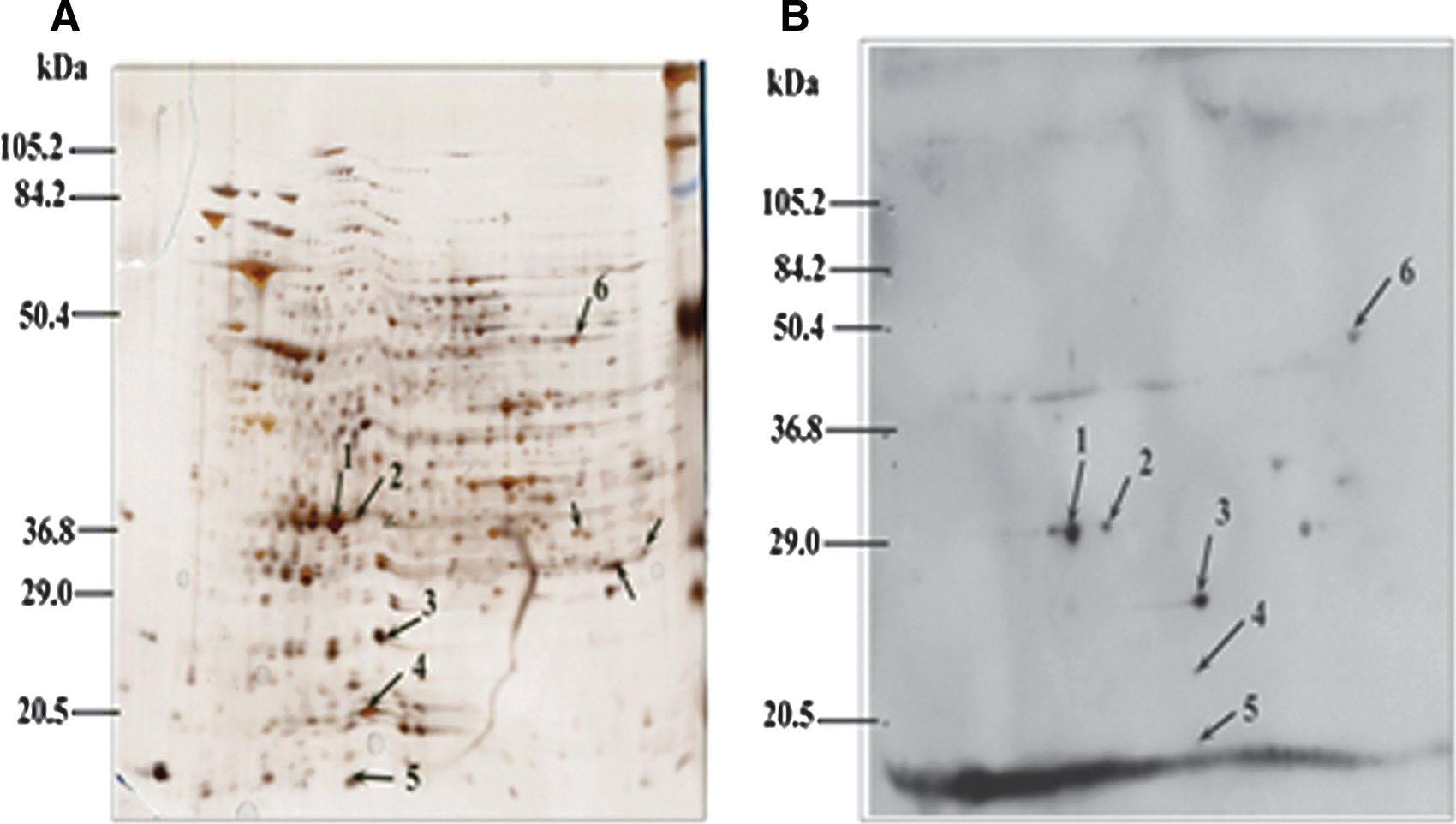
(
The protein spots were excised manually and identified with matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) mass spectrometry.
NCBI, National Center for Biotechnology Information; MS, mass spectometry; MW, molecular weight; pIP, protein isoelectric point.
Discussion
In the present study, we collected molecular and epidemiological data for a co-infection with two arthropod-associated bacteria, O. tsutsugamushi and A. nasoniae. This co-infection was confirmed by two different serological testing methods and molecular-based techniques. We provided molecular evidence of the DNA of these bacteria in a skin biopsy taken 1 week after the onset of disease. The DNA of A. nasoniae was detected with two different primers and probes for qPCR, and we obtained a sequence with a high percentage of similarity with A. nasoniae (GenBank accession no. FN545258). This patient is the only one for which we detect DNA from A. nasoniae among the 154 skin biopsy tested between 2010 and 2011, and 2 patients included our case were positive with specific PCR for O. tsutsugamushi.
We tested the patient's sera by indirect immunofluorescence assay and detected seroconversion with species-specific antibodies against A. nasoniae and O. tsutsugamushi serotype Gilliam at 1 month after the onset of symptoms. These results unquestionably indicate a recent infection, and western blotting confirmed these results. This case may present the first proof of the pathogenicity of the A. nasoniae for humans. Moreover, we identified six immunogenic proteins from A. nasoniae (strain IZ40 isolated from I. ricinus) with polyclonal mouse anti-A. nasoniae antibodies by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (Fig. 4B). Identified immunogenic proteins could be used as the target for serological diagnostic of the infection caused by A. nasoniae.
The serological and molecular data show that our patient was co-infected with A. nasoniae and O. tsutsugamushi. Several cases of co-infection with scrub typhus have been described in the literature and usually involve leptospirosis (Wang et al. 2003, Suttinont et al. 2006, Chen et al. 2007, Lee and Liu 2007). Patients predominantly present with nonspecific symptoms, such as fever, headache, and myalgia, and clinical signs of leptopsirosis, such as jaundice and acute renal dysfunction. In addition, they sometimes present with a cutaneous rash, lymphadenopathy, or an inoculation eschar, favoring a diagnosis of scrub typhus. Co-infection with dengue, influenza, and Japanese encephalitis virus, particularly in patients with undifferentiated febrile illness in Thailand, has also been described (Suttinont et al. 2006).
Reports of co-infection with O. tsutsugamushi and other pathogens in vectors are rare in the literature. The co-infection of O. tsutsugamushi and hantavirus has been described in mites (wild Leptotrombidium scutellare) collected in China (Deng et al. 2008). A. nasoniae has also been found in arthropods, particularly in Araneae, Blattaria, Diptera, Hemiptera, and Hymenoptera (Duron et al. 2008), as well as in whiteflies, triatomine bugs, bees, aphids, hippoboscids, and lice (Novakova et al. 2009). A. nasoniae has also been found in hard ticks of the genera Amblyomma and Dermacentor in North America, and recently, in I. ricinus in Europe (Mediannikov et al. 2012). This may also explained by the presence of A. nasoniae in the larvae of parasitic wasps, Ixodiphagus hookeri, as proved for Wolbachia spp. (Plantard et al. 2012). No data are available concerning the presence of Arsenophonus in hard ticks from other regions, in soft ticks, in mites, or in other acariforms. Co-infection in arthropods is known to exist. Ixodes ticks can harbor Borrelia burgdorferi, the causative agent of Lyme disease, and Anaplasma phagocytophilum, the human granulocytic anaplasmosis agent, along with Rickettsia spp., Babesia spp., and Francisella tularensis (Nieto and Foley 2009, Reis et al. 2010). Arthropods are candidates for co-infection with different pathogens because of their potential exposure to multiple species of microorganisms via feeding on multiple hosts. Many ticks can harbor two or more infectious agents and transmit these pathogens simultaneously (Swanson et al. 2006). To date, no common vectors for both A. nasoniae and O. tsutsugamushi have been identified. The present case, however, may be a co-infection without a simultaneous contamination. The possibility that the A. nasoniae could have been delivered to the patient by a different arthropod, but through the same inoculation site, may not be excluded.
Conclusion
In the present study, the laboratory data for our patient suggest that she had a co-infection with A. nasoniae and O. tsutsugamushi. However, clinically, our patient presented typical symptoms of scrub typhus, including fever, lymphadenitis, and an inoculation eschar that normally occurs between 7–21 days after the initial bite. The implications of A. nasoniae in human pathology remain unclear. A. nasoniae has not been described in mites, but co-infection with different pathogens may occur in the same vector. We have not established any link between A. nasoniae and mites, however, that needed further investigation. Therefore, we will continue to screen for the presence of A. nasoniae in skin from patients bitten by arthropods.
Footnotes
Author Disclosure Statement
No competing financial interests exist