
Abstract
In this study, we aimed to identify and genetically characterize spotted fever group (SFG) rickettsiae in ticks, domestic one-humped camels, and horses from farms and Bedouin communities in southern Israel. A total of 618 ixodid ticks (Hyalomma dromedarii, Hyalomma turanicum, Hyalomma excavatum, and Hyalomma impeltatum) collected from camels and horses, as well as 152 blood samples from 148 camels and four horses were included in the study. Initial screening for rickettsiae was carried out by targeting the gltA gene. Positive samples were further analyzed for rickettsial ompA, 17kDa, ompB, and 16S rRNA genes. Rickettsia aeschlimannii DNA was detected in the blood of three camels and 14 ticks (H. dromedarii, H. turanicum, and H. excavatum). Rickettsia africae was found in six ticks (H. turanicum, H. impeltatum, H. dromedarii, and H. excavatum). In addition, Rickettsia sibirica mongolitimonae was detected in one H. turanicum tick. These findings represent the first autochthonous detection of R. africae in Israel. Previous detections of R. africae in Asia were reported from the Sinai Peninsula (Egypt) and Istanbul, only. Furthermore, we report for the first time the finding of R. aeschlimannii in H. turanicum and H. excavatum ticks, as well as the first identification of R. sibirica mongolitimonae in H. turanicum ticks. The tick species identified to harbor R. africae and other SFG rickettsiae have been reported to occasionally feed on people, and, therefore, physicians should be aware of the possible exposure of local communities and travelers, especially those in contact with camels, to these tick-borne rickettsial pathogens.
Introduction
S
Various animals are susceptible to SFG rickettsial infections and may develop rickettsemia; however, the role of vertebrates as reservoirs of some SFG rickettsiae has yet to be determined (Parola and Raoult 2001). Camels may show a heavy burden of ixodid ticks, and Hyalomma dromedarii is the commonest tick species reported to infest camels (Abdel-Shafy et al. 2012, Fard et al. 2012). Other tick species reported to infest camels include Hyalomma impeltatum, H. marginatum, Hyalomma excavatum, and Hyalomma anatolicum (Abdel-Shafy et al. 2012, Fard et al. 2012). The occurrence of R. aeschlimannii and Rickettsia africae in Hyalomma ticks in camels from Africa and the Sinai Peninsula was recently reported (Morita et al. 2004, Abdel-Shafy et al. 2012). In addition, the presence of a Rickettsia-like organism was described in a dromedary herd in Oman; however, no molecular identification was performed (Wernery et al. 2001). This study aimed to identify and genetically characterize SFG rickettsiae in ticks and domestic animals, mainly camels from Bedouin settlements and farms located in the Dead Sea area and the Negev Desert, Israel.
Materials and Methods
Ticks
During February to September, 2012, 618 ixodid ticks were collected from 68 domestic one-humped camels and two horses from the Negev Desert and Dead Sea area, Israel. The collection sites included locations near the northern Dead Sea close to Jericho (sampled twice; February and May), Arad, Dimona, Avdat (sampled twice; August and September), and Nitzana in the Negev Desert (monthly collection between February to September; Fig. 1). Seven camels were sampled twice on different dates for ectoparasites in the Dead Sea area near Jericho and in Avdat. The ticks collected were kept in vials with 70% ethyl alcohol until analyzed. They were morphologically identified (by K.Y.M. and D.A.A.) using standard taxonomic keys.
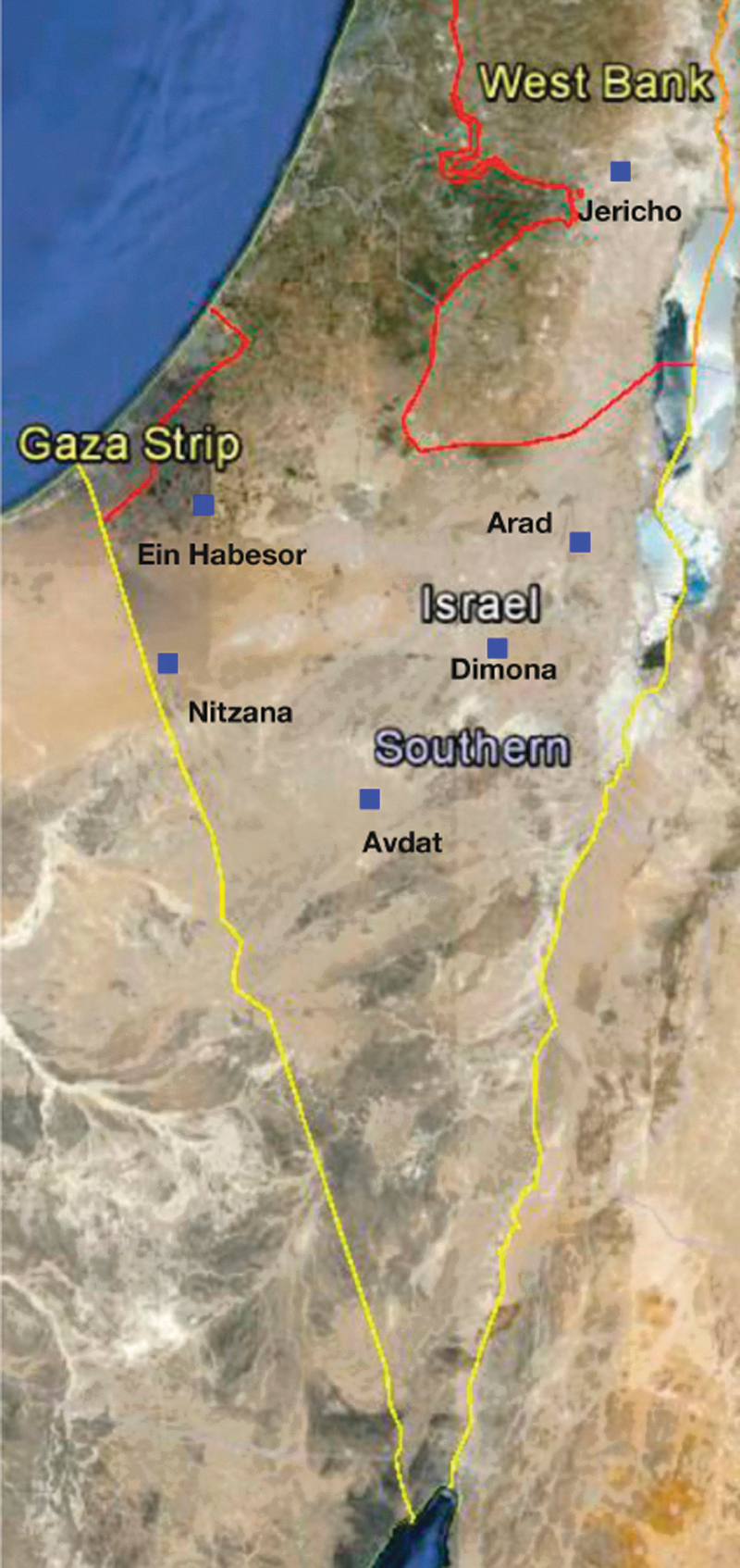
Sample locations marked with small blue rectangles in the Negev Desert and the Death Sea area, Israel, and West Bank. Google Earth (version: 7.0.2.8415) [Software] Google Inc. (2012). Color images available online at
Blood
Blood samples were collected from 44 camels during 2012 from the same locations mentioned above. In addition, samples from 104 camels and four horses collected in 2009 from the same sites and from Ein Habesor in the northern Negev Desert were also used in the study. Blood was drawn from the jugular vein and kept in EDTA vials at −80°C until analyzed further.
DNA extraction, PCR amplification, and sequencing
Ticks were washed individually with phosphate-buffered saline (PBS) and air dried for 15 min on paper. Because most ticks were unengorged, the whole unengorged tick was separately sliced into minute pieces by a sterile scalpel blade. When engorged ticks were included, only the anterior part of the tick was used. Thereafter, they were manually crushed and homogenized in a tube with 50μL of PBS using plastic microtube pestles for 1 min. DNA was extracted from ticks with the Illustra Tissue and Cells Mini Spin kit and from blood using the Illustra Blood Mini Spin kit (GE Healthcare, Little Chalfont, UK), according to the manufacturer's instructions. PCR reactions were performed using the oligonucleotide primers listed in Table 1, and the protocols previously described (Roux and Raoult 2000, Paddock et al. 2004, Kidd et al. 2008, Harrus et al. 2011, Abdel-Shafy et al. 2012, Ogo et al. 2012). For initial detection of Rickettsia species, all DNA samples were screened by a real-time PCR, targeting the citrate synthase gene (gltA) (Harrus et al. 2011). Rickettsial gltA-positive samples were analyzed further, targeting the outer membrane protein A (ompA) gene by conventional and real-time PCR (Kidd et al. 2008, Abdel-Shafy et al. 2012), the 17kDa gene by a nested PCR (Paddock et al. 2004), the 16S ribosomal RNA (16S rRNA) gene (Ogo et al. 2012), and the outer membrane protein B (ompB) gene (Roux and Raoult 2000), both by conventional PCR assays, as previously described (Roux and Raoult 2000, Ogo et al. 2012).
Real-time PCR was carried out using the Rotor Gene 6000 cycler (Corbett Research, Sydney, Australia). Each reaction was performed in 20 μL containing 4 μL of DNA, 1.5 μL of each primer, 0.6 μL of syto9 (Invitrogen, CA), 2.4 μL of double-distilled water, and 10 μL of Maxima Hot Start PCR Master Mix (Thermo Scientific, Loughborough, UK). Conventional PCR assays were run by the Biometra thermocycler version 09/07 (Biomerta, Göttingen, Germany). DNA extracted from R. conorii israelensis- and R. aeschlimannii-positive samples confirmed by sequencing was used as positive controls for the initial screening (gltA) and the further analysis (other genes), respectively. Tick and blood samples negative for rickettsial DNA were used as negative controls, and UltraPure Water (Fisher Biotec, Australia) was used as a nontemplate control.
All positive PCR amplicons were purified using a PCR purification kit (Exo-SAP, NEB; New England Biolabs, Inc. Ipswich, MA), and subsequently sequenced using the same pair of primers described above (Table 1), the BigDye Terminator cycle sequencing chemistry from Applied Biosystems (ABI, Carlsbag, CA), ABI 3700 DNA Analyzer, and the ABI Data collection and Sequence Analysis software. Further sequence analysis was carried out using the ChromasPro version 1.5 (South Brisbane, Australia). Statistical analysis was performed using the Fisher exact two-tailed test, in the WINPEPI program (PEPI-for-Windows, Hebrew University, Jerusalem, Israel).
Results
Ticks
A total of 514 H. dromedarii, 51 Hyalomma turanicum, 51 H. excavatum, and 2 H. impeltatum were identified.
Rickettsiae in ticks
Rickettsial gltA DNA was found in 27 of the 618 ticks, in all localities except for Arad (Table 2). Further analysis revealed at least one other positive detection of R. aeschlimannii ompA, 17kdA, 16S rRNA, or ompB gene fragment, with a sequence identity of 99–100% to GenBank accession numbers JQ691734.1, JN871824.1, NR_026042.1, and AF123705, respectively, in 14 of the 27 gltA-positive samples. The ompA amplicons were found in two H. dromedarii, 10 H. turanicum, and two H. excavatum. All of the positive ticks were collected from nine camels, except for one H. excavatum tick, which was collected from a horse.
Rickettsia spp. determined according to ompA (short fragment) sequence identity. R. aeschlimannii accession number JQ691734.1, sequence identity of 100%. R. africae accession number JF700254.1, sequence identity of 99–100%. R. sibirica mongolitimonae accession number JF700255.1, sequence identity of 99%.
SFGR, spotted fever group rickettsiae.
OmpA sequences, 99–100% similar to R. africae (GenBank accession number: JF700254.1), were detected in six ticks, two H. turanicum, two H. impeltatum, one H. dromedarii, and one H. excavatum. The positive H. dromedarii tick was collected from a horse; the rest were collected from four different camels except for one H. turanicum, which was an active questing tick. Finally, DNA extracted from an H. turanicum tick collected from a camel revealed amplicons 99–100% similar to R. sibirica mongolitimonae ompA and 16S rRNA gene fragments (GenBank accession numbers DQ097082.1 and HM050271.1, respectively) (Tables 2 and 3).
Short fragment.
Long fragment.
C, camel; HD, Hyalomma dromedarii; HT, H. turanicum; HE, H. excavatum; HI, H. impeltatum.
Sequencing of the gltA fragment performed for six amplicons that were negative for all other targeted genes shared 99–100% sequence identity with a large number of Rickettsia species. The prevalence of rickettsial gltA DNA in H. turanicum was significantly higher than the prevalence of infection in H. dromedarii (14 of 51 compared with eight of 514 ticks, respectively; p<0.001).
In the Dead Sea area, three H. turanicum ticks out of a total of 40 ticks (34 H. dromedarii and six H. turanicum) collected from two camels in May contained SFG rickettsial DNA; however, none of the ticks (31 H. dromedarii and one H. turanicum) collected from the same animals 3 months earlier were positive. One H. turanicum tick from a camel collected in Avdat in September, 2012, contained rickettsial DNA; nevertheless, ticks of the same species collected from the same animal 1 month before were negative. The molecular prevalence of SFG rickettsiae in ticks from the Dead Sea area was significantly higher in May compared to February, 2012 (nine of 93 compared with one of 173 ticks, respectively; p=0.001). On the contrary, no significant differences were noticed in the prevalence in Avdat between August and September 2012 (four of 135 and three of 45 ticks, respectively; p=0.375).
Rickettsiae in blood
Thirty-two camel samples were positive for rickettsial gltA gene fragment. Out of 15 camels, which were sampled twice (once in 2009 and again in 2012), two were positive for rickettsial gltA when examined in 2009 but negative for the same gene fragment when resampled and tested in 2012. One other camel was negative for this screening test in 2009 but was found positive during sampling in 2012. When positive gltA samples were further analyzed, three samples were found 99–100% similar to R. aeschlimannii ompA DNA (short fragment [Table 1]; GenBank accession number: JQ691734.1) and were detected in camel samples collected in 2009 from Arad, Dimona, and Avdat (Table 3). All blood samples were negative for the 17kDa gene, the 16S rRNA gene, and the ompB gene fragments.
Discussion
In this study, R. africae was detected in H. impeltatum, H. dromedarii, H. turanicum, and H. excavatum ticks. This finding represents the first autochthonous detection of this rickettsia in Israel. The only previous detections of R. africae in Asia were reported in ticks collected from camels and cows from the Sinai Peninsula (Egypt) (Abdel-Shafy et al. 2012) and in ticks removed from humans in Istanbul, Turkey (Gargili et al. 2012). R. africae is the agent of human African tick bite fever, a disease characterized by multiple inoculation eschars in addition to other typical symptoms associated with SFG rickettsioses (Parola et al. 2005). Travelers visiting southern Africa during the summer are at risk of acquiring the disease (Parola et al. 2005, Wieten et al. 2011). In South Africa, R. africae is transmitted mainly by Amblyomma hebraeum. The rickettsia was also detected in Amblyomma lepidum and Amblyomma variegatum in northern and central Africa (Parola et al. 2001, Morita et al. 2004, Parola et al. 2005). However, it is important to note that other tick genera may carry this pathogen, as reported from Egypt, Turkey, Liberia, and Guinea (Abdel-Shafy et al. 2012, Gargili et al. 2012, Mediannikov et al. 2012). R. africae can be considered as an emerging pathogen in Israel, because it was not recognized nor identified previously in this country. The possibility that this pathogen was present in Israel but not detected before could be explained by the fact that no molecular studies for Rickettsia have been carried out in some important tick and animal populations, nor in every suspected SFG infection in humans. However, it is likely that R. africae was introduced to Israel from Egypt, through the Sinai desert, via ticks carried by camels crossing the border to Israel.
The finding of R. aeschlimannii and R. africae in ticks collected from camels in Israel is corroborated by previous findings from Egypt (Abdel-Shafy et al. 2012). Some of the ompA PCR amplicons found in our study were similar to R. aeschlimannii strain EgyRickHimp-El-Arish-17 and R. africae strains EgyRickHmm-Qalet El-Nakhl-2/9, from the Sinai Peninsula. R.africae was detected in the present study in H. excavatum, H. turanicum, H. dromedarii, and H. impeltatum. The latter two tick species were also found to harbor R. africae in Egypt, thus the importation of this Rickettsia into Israel by animals parasitized with Hyalomma ticks from the Sinai Peninsula is a plausible explanation.
R. africae could be an example of a tick-borne disease imported to another region by livestock trade, like the occurrence of SFG rickettsioses associated with this Rickettsia in the West Indies, after cattle parasitized with A. variegatum were imported from Senegal in the early 1800s (Kelly 2006). Another possible route is the carriage of infected ticks by migrating birds, because they act as transport carriers of many species of the Ixodidae family and therefore as long-distance vectors for several pathogens including Rickettsiae spp., like R. aeschlimannii (Movila et al. 2012). It should be noted that Israel is located on the main route of bird migration from Africa to Europe and Asia, thus this route of introduction seems possible (Harrus et al. 2011).
We report here the first detection of R. aeschlimannii in H. turanicum and H. excavatum ticks as well as the first identification of R. sibirica mongolitimonae in H. turanicum. R. aeschlimannii was first characterized as a new SFG Rickettsia after its isolation from H. marginatum in Morocco in 1997 (Beati et al. 1997). Thereafter, two human cases associated with R. aeschlimannii were reported from travelers returning from Africa to Europe with symptoms suggestive of SFG rickettsioses (Parola et al. 2005). H. marginatum is considered to be the main reservoir of R. aeschlimannii (Parola et al. 2005, Keysary et al. 2011, Abdel-Shafy et al. 2012, Gargili et al. 2012). However, R. aeschlimannii was detected in several other species of ticks, including Hyalomma rufipes in Niger and Mali (Parola et al. 2001), H. dromedarii in Sudan (Morita et al. 2004), Hyalomma aegyptium and Rhipicephalus bursa in Turkey (Gargili et al. 2012), H. scupense in Israel (Keysary et al., 2011), and H. impeltatum in Egypt (Abdel-Shafy et al. 2012). This study reports the first detection of R. aeschlimannii in ticks from Israeli camels and horses, and the first identification of this rickettsia in H. turanicum and H. excavatum. Although R. aeschlimannii DNA was identified in three camels, no correlation between SFG rickettsiae in ticks and their hosting camels was found, due to the fact that the positive blood samples were collected in 2009 without simultaneous tick collection. Therefore, further investigation is necessary to study the role of camels in the transmission of SFG rickettsioses.
R. sibirica mongolitimonae, the etiologic agent of lymphangitis-associated rickettsiosis, is clinically characterized by multiple eschars and lymphangitis (Fournier et al. 2005). It was associated with human cases in Europe (Psaroulaki et al. 2005, de Sousa et al. 2006) and South Africa (Pretorius and Birtles 2004) and was first isolated from a Hyalomma asiaticum from Mongolia in 1993 (Yu et al. 1993), and subsequently detected in several Hyalomma and Rhipicephalus species (Parola et al. 2001, de Sousa et al. 2006, Harrus et al. 2011). To the best of our knowledge, this is the first report of R. sibirica mongolitimonae in H. turanicum ticks.
High temperatures and humidity have a great impact on the ecology of ticks, their establishment in new niches, and thereby on the incidence of tick-borne diseases in new geographic areas (Harrus and Baneth 2005). A higher prevalence of SFG rickettsiae was found in ticks from the Dead Sea in May compared to February. This is probably due to a warmer climate during May in the study area, a period when the detected tick species are more active. High temperatures were previously associated with increased prevalence and diversity of ticks in camels (Fard et al. 2012), possibly explaining the findings in our study.
Six DNA samples extracted from ticks and 29 DNA samples extracted from blood were positive only for rickettsial gltA DNA but not for other rickettsial genes. Several Rickettsia spp. are known to exist as endosymbionts in ixodid ticks (Rounds et al. 2012). Endosymbionts were shown not to express certain proteins associated with pathogenicity, like the ompA gene. Therefore, the absence of the latter gene may suggest that they were endosymbiont rickettsiae rather than pathogenic ones (Felsheim et al. 2009).
The number of yearly recorded cases of rickettsial infection in humans in Israel is low and most probably underreported. According to the Israeli Ministry of Health website, only 11–13 patients suffering from rickettsiosis were documented in 2011–2012 (retrieved on November, 2012,
Some populations, such as Bedouin shepherds and farmers, are in continuous exposure to ticks parasitizing their domestic animals and consequently at risk of contracting rickettsial infections. Therefore, physicians should be aware of the possible exposure to these pathogens in the Negev Desert and Dead Sea areas of Israel, when travelers, Bedouins, farmers, or other patients present with fever or other clinical signs compatible with rickettsioses. Moreover, most of the serological tests carried in Israeli hospitals use R. conorii as an antigen when a rickettsial disease is suspected, and no discrimination between the different SFG rickettsiae is carried out. Thus, molecular identification is vital in identifying the specific SFG Rickettsia spp. involved.
Footnotes
Acknowledgment
This study was funded by The Ministry of Foreign Affairs, The Hague, The Netherlands, through a grant to the Dutch Friends of the Hebrew University (NVHU).
Author Disclosure Statement
No competing financial interests exist.