
Abstract
Bartonella spp. are fastidious, Gram-negative bacilli that cause a wide spectrum of diseases in humans. Most Bartonella spp. have adapted to a specific host, generally a domestic or wild mammal. Dromedary camels (Camelus dromedarius) have become a focus of growing public-health interest because they have been identified as a reservoir host for the Middle East respiratory syndrome coronavirus. Nevertheless, data on camel zoonoses are limited. We aimed to study the occurrence of Bartonella bacteremia among dromedaries in Israel. Nine of 51 (17.6%) camels were found to be bacteremic with Bartonella spp.; bacteremia levels ranged from five to >1000 colony-forming units/mL. Phylogenetic reconstruction based on the concatenated sequences of gltA and rpoB genes demonstrated that the dromedary Bartonella isolates are closely related to other ruminant-derived Bartonella spp., with B. bovis being the nearest relative. Using electron microscopy, the novel isolates were shown to be flagellated, whereas B. bovis is nonflagellated. Sequence comparisons analysis of the housekeeping genes ftsZ, ribC, and groEL showed the highest homology to B. chomelii, B. capreoli, and B. birtlesii, respectively. Sequence analysis of the gltA and rpoB revealed ∼96% identity to B. bovis, a previously suggested cutoff value for sequence-based differentiation of Bartonella spp., suggesting that this approach does not have sufficient discriminatory power for differentiating ruminant-related Bartonella spp. A comprehensive multilocus sequence typing (MLST) analysis based on nine genetic loci (gltA, rpoB, ftsZ, internal transcribed spacer (ITS), 16S rRNA, ribC, groEL, nuoG, and SsrA) identified seven sequence types of the new dromedary isolates. This is the first description of a Bartonella sp. from camelids. On the basis of a distinct reservoir and ecological niche, sequence analyses, and expression of flagella, we designate these isolates as a novel Bartonella sp. named Bartonella dromedarii sp. nov. Further studies are required to explore its zoonotic potential.
Introduction
Bartonella
Several Bartonella spp. have been described in cattle and wild ruminants, such as roe deer, mule deer, elk, and water buffalo (Chang et al. 2000, Dehio et al. 2001, Bermond et al. 2002, Bai et al. 2011, Bai et al. 2013). Cattle comprise the main reservoir of B. bovis and B. chomelii (Bermond et al. 2002, Maillard et al. 2004), whereas B. schoenbuchensis and B. capreoli were first described in roe deer (Dehio et al. 2001, Bermond et al. 2002). Additionally, B. melophagi was first isolated from commercial sources of sheep blood, and its DNA was detected in sheep ked (Bemis and Kania 2007, Kumsa et al. 2014). The clinical relevance of these species to humans is unclear, although Candidatus Bartonella melophagi was isolated from the blood of two female patients (Maggi et al. 2009). Bartonella infection in ruminants is usually subclinical. However, in a few cases, B. bovis was identified as a cause of prolonged bacteremia and bovine endocarditis (Maillard et al. 2007, Erol et al. 2013).
Dromedary camels (Camelus dromedarius) have a close association with humans; in addition to being employed for transportation of people and goods in some camel-rearing countries and a valuable resource for milk, meat, and wool, camels are also recreational animals used for camel racing and tourism. Camels have been uncommonly implicated as a source for zoonoses. In fact, apart from camel brucellosis, camel-to-human transmission of zoonotic agents is either rare or unsubstantiated. Nonetheless, camels have been proposed as the reservoir for the newly described Middle East respiratory syndrome coronavirus (Alagaili et al. 2014, Haagmans et al. 2014). Given the apparent growing scientific and public health interest in camels as potential zoonotic hosts under a “One Health” approach, we decided to examine the prevalence of Bartonella spp. in domestic dromedary camels from Israel. In this study, we describe the first known Bartonella sp. from camelids. Their potential role as reservoir for human bartonellosis warrants further investigation.
Materials and Methods
Animals and specimen collection
During May of 2013, a total of 51 dromedary camels from three camel farms located in central and southern Israel were studied (herd A, n=9; herd B, n=15 and herd C, n=27). Dromedary blood samples (2–3 mL of whole blood in EDTA-coated tubes) were obtained by jugular vein puncture according to standard veterinary procedures by trained personnel. Samples were kept at −80°C until processed. Data on gender, age group (<10 years, 10–20 years, >20 years old), and the presence of ectoparasites were recorded for each animal. This study was approved ethically by the Tel Aviv Sourasky Medical Center Institutional Animal Care and Use Committee (study no. 18-6-13).
Culture conditions and Bartonella spp. isolation
Dromedary blood samples were thawed, plated in duplicates on fresh chocolate agar plates (Novamed, Jerusalem, Israel), and incubated at 37°C in a humid 5% CO2 atmosphere for 8 weeks or until growth. Plates were inspected twice weekly for growth. Putative identification of Bartonella sp. was based on Gram stain and morphological examination of the colonies. Quantitative evaluation of the number of colonies and their morphology were recorded. Bacterial stocks were prepared in brain heart infusion broth (Novamed, Jerusalem, Israel) and 15% glycerol vol/vol (Sigma Aldrich, Rehovot, Israel) and kept at −80°C.
DNA extraction
Bacterial colonies were scraped off chocolate agar plates, washed once, and resuspended in 200 μL of sterile 1× phosphate-buffered saline (PBS; Sigma Aldrich, Rehovot, Israel). Genomic DNA was extracted using the QIAamp DNA Mini Kit (QIAGEN, Valencia, CA) according to the manufacturer's instructions.
PCR and sequencing
Conventional PCR was performed with Q5® or Phusion® High-Fidelity DNA Polymerase (New England Biolabs, Hitchin, UK) according to the manufacturer's protocol. The primers (Sigma Aldrich, Rehovot, Israel) used in this study are listed in Table 1. One or 2 μL of genomic DNA preparations of Bartonella isolates were used as template. PCR products were analyzed by gel electrophoresis and purified with Illustra® ExoProStar 1-Step Kit (GE Healthcare, Buckinghamshire, UK). Sequencing was performed in the DNA Sequencing Unit at the G.S. Wise Faculty of Life Sciences, Tel Aviv University. Nucleotide sequences were submitted to a BLAST search in the National Center for Biotechnology Information (NCBI) GenBank database with default parameters, while excluding uncultured or environmental sample sequences.
ITS, internal transcribed spacer.
Phylogenetic reconstruction
Phylogenetic reconstruction was conducted using MEGA software version 5.1 (Tamura et al. 2011). Alignments were performed with the MUSCLE algorithm (Edgar 2004) with default parameters. Phylogenetic relations were inferred using the neighbor-joining method (Saitou and Nei 1987). The evolutionary distances were computed using the Jukes–Cantor model (Jukes and Cantor 1969). The partial gltA and rpoB sequences of the novel dromedary isolates were included in this analysis, along with the respective nucleotide sequences of valid Bartonella species and Candidatus B. melophagi. Bootstrap analysis (Felsenstein 1985) was carried out on 1000 replications of the dataset.
Biochemical phenotypes
The enzymatic profiles of Bartonella isolates were obtained using Vitek®2 (bioMérieux, Durham, NC), an automated bacterial identification system using GN ID and NH ID cards (product numbers 21341 and 21346, respectively). Oxidase and catalase tests were performed using standard bacteriological procedures.
Electron microscopy
Bacteria were submitted to electron microscopy, as described elsewhere (Bermond et al. 2002), with minor modifications. Briefly, bacteria were scraped from chocolate agar plates, suspended in sterile 1× PBS (Bio-Lab, Jerusalem, Israel), and adsorbed to Formvar-coated copper grids. Grids were then stained with 1% (wt/vol) uranyl acetate and air dried. Negative-stained micrographs of cell morphology were analyzed using a transmission electron microscope (TEM) Tecnai 12 (Phillips, Eindhoven, The Netherlands) at 100 kV equipped with MegaView II CCD camera and Analysis® v. 3.0 software (Soft Imaging System GmbH, Münstar, Germany).
Multilocus sequence typing
Sequence data of dromedary Bartonella isolates was obtained for nine genetic loci listed in Table 1, similarly to the multilocus sequence typing (MLST) scheme previously defined for B. bovis (Bai et al. 2013). For determination of allelic variations, nucleotide sequences were aligned using Clustal Omega multiple sequence alignment program with default parameters (
Nucleotide sequence accession numbers
Nucleotide sequences were deposited in the NCBI GenBank database with the accession numbers listed in Table 2.
ITS, internal transcribed spacer.
Results
Prevalence of Bartonella sp. bacteremia
Fifty-one animals were included in this study; 41% were <10 years old, 53% 10–20 years old, and 6% >20 years old. Thirty-three (65%) of the 51 camels were females. Nine of 51 (18%) camels sampled were found to be bacteremic with Bartonella sp., with concentrations ranging from five to >1000 colony-forming units (CFU)/mL (median=63). Round, small (3–5 mm in diameter), homogeneous, opaque, grey-white bacterial colonies appeared after 14.5±1.6 days of incubation. Subcultures growth time was approximately 7 days under the same conditions.
Molecular identification and phylogenetic analysis
Sequence comparison analysis of the partial gltA and rpoB sequences of the dromedary Bartonella isolates demonstrated ∼96% identity to B. bovis for each of the amplified sequences, contrary to 100% identity demonstrated for a recently characterized bovine B. bovis isolate from Israel (Rudoler et al. 2014), using the same targets. Given the considerable difference (4%) between the gltA and rpoB sequences of the dromedary isolates and B. bovis and the resulting homologies, which are at the low range of the identity values proposed for species differentiation (La Scola et al. 2003), we decided to expand our analysis. Sequence comparison analysis of the housekeeping genes ftsZ, ribC, and groEL showed the highest homology to B. chomelii, B. capreoli, and B. birtlesii, respectively (Table 3). These data demonstrate that the dromedary isolates are not B. bovis.
NADH, nicotinamide adenine dinucleotide; ITS, internal transcribed spacer.
A phylogram based on the concatenated sequences of gltA and rpoB demonstrated that all dromedary Bartonella isolates comprise a distinct monophyletic group with B. bovis (bootstrap test=81%) (Fig. 1). As expected, additional ruminant-associated Bartonella spp. (B. capreoli, B. schoenbuchensis, B. chomelii, and Candidatus B. melophagi) were clustered together in the adjacent sister clade, which suggests a common evolutionary ancestor to Bartonella spp. that infect ruminants (bootstrap test=100%). This phylogenetic analysis clearly indicates that the dromedary isolates found in this study are closely related to ruminant-derived Bartonella spp., and B. bovis is the nearest evolutionary relative.
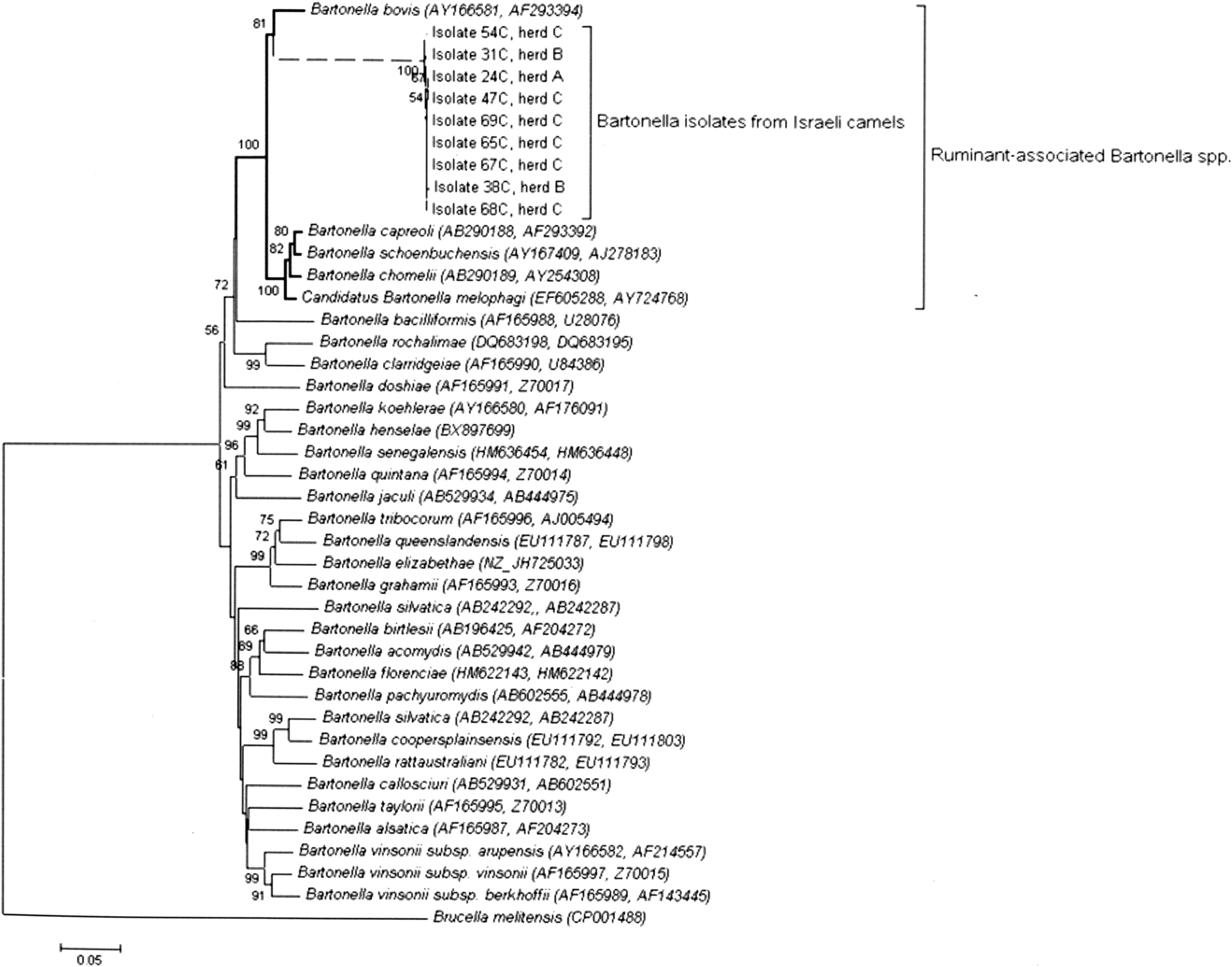
A phylogenetic tree highlighting the position of Bartonella isolates form Israeli dromedary camels relative to other type strains within the genus Bartonella. Concatenated gltA and rpoB sequences were aligned using MUSCLE (Edgar 2004), and phylogenetic interferences were obtained using the neighbor-joining method (Saitou and Nei 1987). The optimal tree with the sum of branch length=1.99 is shown. GenBank accession numbers are indicated in parentheses as (rpoB, gltA). The tree was rooted using Brucella melitensis as an outgroup. Numbers at the nodes are the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) (Felsenstein 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. Branches drawn with a dashed line represent the dromedary isolates described in this study. The evolutionary distances were computed using the Jukes–Cantor method (Jukes and Cantor 1969), and the scale bar indicates 10% nucleotide sequence divergence. The analysis involved 42 nucleotide sequences. There were a total of 1093 positions in the final dataset. All evolutionary analyses were conducted in MEGA5 (Tamura et al. 2011).
Dromedary Bartonella isolates are flagellated
Electron micrographs of three representative dromedary isolates (one from each herd) (Fig. 2a–c), as well as a B. bovis isolate from Israeli beef cattle (Rudoler et al. 2014) (Fig. 2d) were obtained. All isolates submitted to TEM examination were grown and harvested under the same conditions. The bacterial cells were found to be rod-shaped and approximately 850 nM in length and 450 nM in width. As expected, a double-layer membrane characteristic of Gram-negative bacteria was also observed. All three dromedary isolates were found to carry unipolar flagella in contrast to the bovine B. bovis isolate, which was nonflagellated. A B. bovis strain previously isolated from a French cow was also shown to be nonflagellated (Bermond et al. 2002), in agreement with our observation. Finally, wet-mount slide tests confirmed that the dromedary Bartonella isolates are motile, in contrast to the nonflagellated B. bovis.
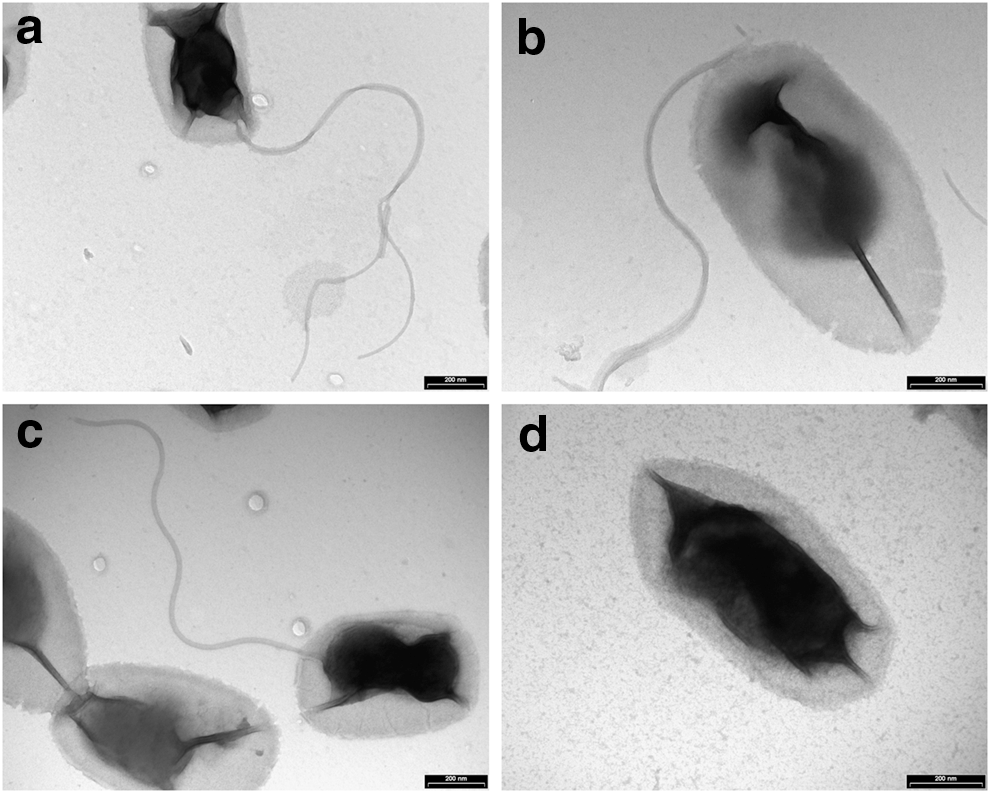
Dromedary Bartonella isolates are flagellated. Transmission electron micrographs of negatively stained cells of Bartonella isolates 24C (
Biochemical phenotypes
All dromedary isolates were initially characterized as oxidase (−), catalase (−), Gram-negative bacilli. Using the Vitek® 2 system, representative isolates were found to have amino acid arylamidase activity with the following amino acids: Arginine, phenylalanine, proline, glycine, tyrosine, and leucine. One of the isolates was also shown to have lysine arylamidase activity; however, they did not have alanine-phenylalanine-proline arylamidase activity. The bacteria lacked the ability to hydrolyze mono- or disaccharides (i.e., glucose, galactose, mannose, trehalose, maltose, sucrose, tagatose, ribose, and xylose), had no urease activity, and were unable to hydrolyze N-acetyl-β-
Multilocus sequence typing
To further characterize the novel dromedary Bartonella isolates and determine the prevalent sequence types (STs), a MLST analysis was performed. Twenty-six of 6187 (0.42%) nucleotide positions analyzed were variable, suggesting close genetic relatedness among the examined isolates. Each dromedary isolate was characterized by a series of nine integers that correspond to the alleles at the internal fragments of nine housekeeping loci, and a total of seven STs were assigned. Of note, the partial gltA sequences of the dromedary isolates were identical. ST1 was the prevalent ST, and the only ST found in more than one camel farm (Table 4).
ITS, internal transcribed spacer; ST, sequence type.
Tick infestation
High tick infestation rate (80%) was found in herd B, as opposed to the other two herds not infested with ectoparasites, probably due to prior anti-ectoparasite treatment. Eighteen ticks, representing one or two ticks from each of the 11 camels out of 12 infested animals, were classified by an entomologist as Hyalomma spp. Three ticks were males and the rest were females at different stages of feeding.
Discussion
In this study, we isolated and characterized a novel Bartonella sp. from dromedary camels inhabiting camel farms located in the Negev desert and central Israel. To the best of our knowledge, this is the first report of a Bartonella sp. from camels. Bartonella sp. bacteremia was found in all of the camel herds examined with an overall prevalence of 18%. Bartonella dromedary isolates displayed phenotypic traits characteristic of the genus Bartonella: They were isolated from blood, grew slowly (14 days for primary isolation and 7 days for subcultures), and produced small, round, homogeneous, grey-white colonies on chocolate agar plates, with straight and slender Gram-negative rods. These isolates can be briefly described as small, aerobic, fastidious, oxidase (−) and catalase (−), Gram-negative bacteria, which are nonfermenters and biochemically inert except for the production of peptidases. Such enzymatic profiles are characteristic of the Bartonella genus (Bermond et al. 2002, Maillard et al. 2004); consequently, biochemical profiles cannot be used routinely for the differentiation of Bartonella spp.
We initially applied a straightforward and well-known molecular approach for the identification of newly encountered Bartonella isolates, as was suggested by La Scola and colleagues (La Scola et al. 2003). DNA sequence comparisons of the partial gene fragments of two Bartonella housekeeping genes (gltA and rpoB) of the dromedary isolates revealed 96% identity to B. bovis sequences for each one of the targets (Table 3); the latter were chosen for this analysis due to their good discriminatory power. According to La Scola's criteria, a newly encountered Bartonella isolate should be considered a new species if a 327-bp gltA fragment shares less than 96.0% sequence similarity and a 825-bp rpoB fragment shares less than 95.4% sequence similarity with those of the validated species (La Scola et al. 2003). Based exclusively on these criteria, the dromedary isolates described here should be considered as B. bovis. However, a more comprehensive sequence comparisons analysis based on nine genetic loci suggested that the dromedary isolates represent a novel Bartonella sp., which is evidently different from, although evolutionarily related to, B. bovis (Table 3 and Fig. 1).
We suggest that a sequence-based approach for the differentiation of Bartonella spp. that utilizes the partial sequences of gltA and rpoB does not have sufficient discriminatory power in relation to ruminant-related Bartonella spp. If that is the case, then the La Scola paradigm should be revised. Accordingly, to fully differentiate members of this group, more robust methods should be taken, such as sequence determination of multiple targets with high discriminatory power or whole-genome sequence comparisons. A MLST scheme, on the basis of nine genetic loci, originally developed to compare genetic variants of B. bovis strains (Bai et al. 2013), was modified in our study and identified seven STs of the new dromedary isolates (Table 4). This MLST scheme may serve as a future reference for comparing camel-associated Bartonella isolates.
Dromedary Bartonella isolates were also found to be flagellated (Fig. 2). Motility is not a known trait of B. bovis, as a previously reported bovine B. bovis isolate from France (Bermond et al. 2002) as well as a B. bovis isolate from cattle in Israel (Rudoler et al. 2014) were shown to be nonflagellated. Thus far, ruminant-derived Bartonella spp. B. chomelii, B. schoenbuchensis, B. capreoli, and Candidatus B. melophagi were shown to carry flagella (Dehio et al. 2001, Bermond et al. 2002, Maillard et al. 2004, Maggi et al. 2009). B. bovis is an exception, because it is the only nonflagellated member among the species comprising lineage II of the phylogenetic tree of the genus Bartonella (Fig. 1 and Harms and Dehio 2012). The dromedary Bartonella isolates were found to be closely related to all of these species, because they share the same evolutionary lineage.
Camels have adapted to desert life, which presents temperature extremes and a scarce supply of food and water. Seasonal variations and the hydration/dehydration status of the camel were shown to have significant effects on numerous hematological indices, endocrine function, as well as blood minerals and metabolites concentrations of dromedaries. In addition, the camel's body temperature fluctuates dramatically (34°C–42°C) during the day to minimize water loss (Amin et al. 2007, Ouajd and Barhoumi 2009). It is reasonable to assume that the unique physiology of these mammals required host-specific adaptations of a Bartonella sp. infecting camels. One may also speculate that expression of flagella in the novel dromedary Bartonella isolates is host dependent and tightly regulated by a yet unknown environmental signal(s). In a previous report, a B. melophagi isolate from a sheep ked was shown to be flagellated, whereas no flagella were observed by TEM of a Candidatus B. melophagi human isolate, identified on the basis of sequence homology of the rpoB, gltA, 16S rRNA, and internal transcribed spacer (ITS) fragments (99.2%, 97.9%, 99.7%, and 99.2% similarity, respectively) (Maggi et al. 2009). This is an example of strain variation within a Bartonella sp. that is manifested by differential phenotypic expression of flagella. This phenomenon might be strain or host dependent and warrants further investigation.
Conclusions
To conclude, we show for the first time that dromedary camels may serve as natural hosts for a Bartonella sp. On the basis of distinct reservoir host and ecological niche, sequence-based analyses, and expression of flagella, we designate dromedary Bartonella isolates described herein as a novel Bartonella sp. named Bartonella dromedarii sp. nov. after its host, and assign isolate 24C as the type strain of this species.
The notable prevalence of B. dromedarii bacteremia found in Israeli dromedary camels combined with the close human-to-camel contact and observed abundance of ticks constitute a potential risk for zoonotic transmission. Continuous surveillance of Bartonella infection in camels and in persons who come in close and frequent contact with camels (e.g., camel handlers) are needed to understand the potential role of B. dromedarii as a zoonotic agent.
Footnotes
Acknowledgments
Electron microscopy work was done at the Bio-Imaging Unit, the Alexander Silberman Institute of Life Science, the Hebrew University, Jerusalem, Israel. The authors thank Dr. Yuval Gottlieb-Dror from Koret School of Veterinary Medicine, the Hebrew University of Jerusalem for morphological classification of ticks.
Author Disclosure Statement
No competing financial interests exist.