
Abstract
During arbovirus surveillance in Ixodes ricinus ticks in South Ukraine, two strains of Tribeč virus were isolated using an in vivo method and characterized using the complement fixation text (CFT), hemagglutination assay (HA), electron microscopy, and molecular methods. Both strains replicated well in the BHK-21, PEC, and Vero cell lines and demonstrated 95% nucleotide identity in their VP3 sequences compared with the reference VP3 sequence for Tribeč virus. These two strains of Tribeč virus were named Tr35 and Tr19. Also, segment reassortment involving Tr35, Tr19, TRBV, and LIPV strains was shown.
Introduction
H
Over the past 5 years, interest in orbiviruses has been increasing, and some publications have focused on the characterization of novel orbiviruses, as well as formally known, although poorly characterized, strains and species of the Orbivirus genus (Cowled et al. 2009, Belaganahalli et al. 2011, Belaganahalli et al. 2012, Kapoor et al. 2013, Cooper et al. 2014). To contribute to the study of orbiviruses, we report the study of two strains of TRBV, Tr35 and Tr19, isolated in southern Ukraine.
Materials and Methods
Sampling
During an arbovirus surveillance and control program, ixodid ticks were collected in South Ukraine, and two viral strains were isolated. Strain Tr35 was isolated from 75 questing males of Ixodes ricinus ticks collected by flagging (0.7×1.1 m) from vegetation near Dibrovka village (49°10′N, 30°47′E) in the Kotovsky district of the Odessa region in 1988.
Strain Tr19 was isolated from three female I. ricinus ticks collected from a dog in the Serbino hole (47°56′N, 29′37′E) in the Balta district of the Odessa region in 2008 (Fig. 1). The I. ricinus identity and sex of the ticks were determined based on morphology following the recommendations of Dr. Filippova (1977). All of the ticks were stored alive in a wet chamber until evaluation.
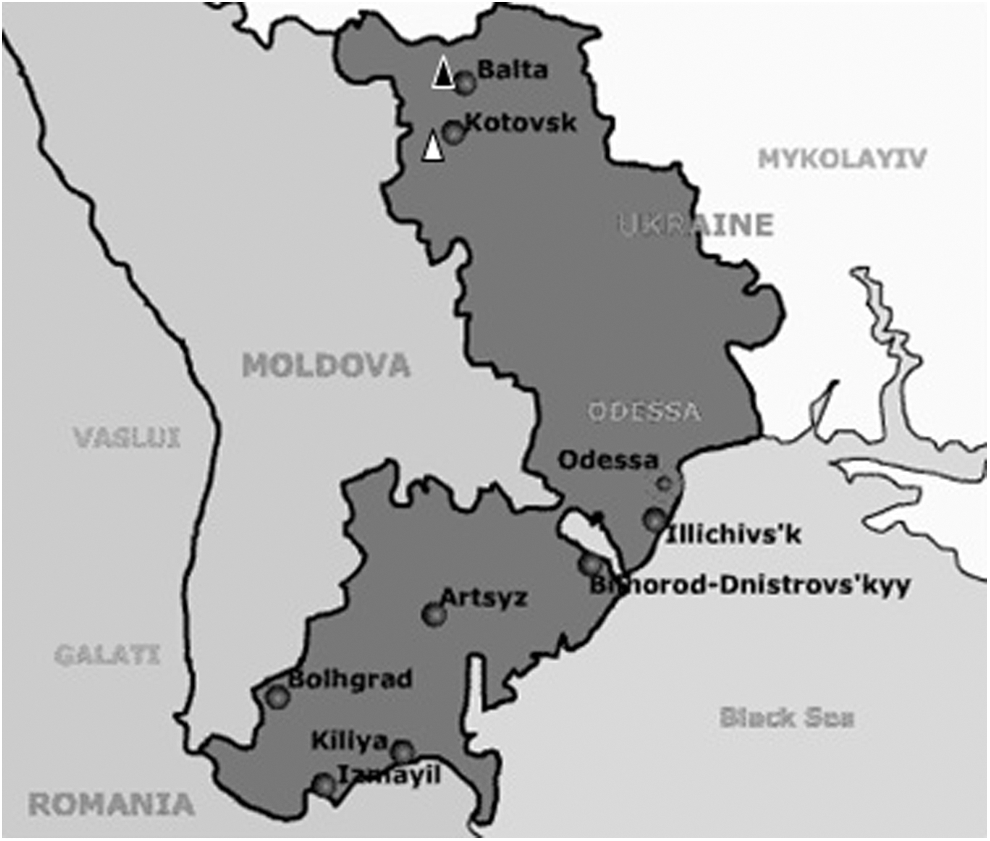
Map of the Odessa region.
Virus isolation and propagation
Virus isolation and propagation were performed using an in vivo method. Briefly, whole tick bodies were pooled and homogenized in Hanks' Buffered Salt Solution (HBSS) (Maniatis et al. 1982). This 10% tick suspension was centrifuged at 2000 rpm for 10 min, and 10–20 μL of the supernatant was inoculated into newborn white mice using the intracerebral route.
The inoculated mice were observed for 21 days. Mice with specific signs of neurotropic disorders (anorexia, flaccidity, paresis, plegia) were euthanized, and the brain was extracted. A 10% brain suspension was prepared and filtered through a 0.22-μm filter (Millipore) to exclude bacterial and protozoal agents, and a second inoculation was performed for virus propagation.
All of the procedures involving mice were performed with the approval of the institutional animal care and use committees.
Virus identification
Identification of the Tr35 and Tr19 strains was performed using the hemagglutination assay (HA) and the complement fixation test (CFT) using acetone-extracted antigen from the mouse brain, as described by Clarke and Casals (1958). Hemagglutination testing was performed at a range of pH values from 6.8 to 7.2 with goose, sheep, and human type O erythrocytes.
The CFT for identifying the Tr35 strain was performed using ascites fluids against KEMV, TRBV, Chenuda virus (CNUV), and Colorado tick fever virus (CTFV). Ascites fluids were produced from mouse ascites sarcoma 180/TG (Hermann and Engle 1958). Ascites fluid against the Tr35 strain was used as a positive control, and serum from an uninfected mouse was used as a negative control.
The CFT for identifying the Tr19 strain was performed using immune sera against TRBV (strain Tr35), Uukuniemi virus (UUKV), and tick-borne encephalitis virus (TBEV). Immune serum against the Tr19 strain was used as a positive control, and serum from an uninfected mouse was used as a negative control.
Ascites fluids and immune sera were titered from 1:8 to 1:256 with two-fold dilutions. The reactions were categorized as 0, 1, 2, 3, and 4, where 0=a complete lack of fixation (total hemolysis), 1=the majority of erythrocytes were hemolyzed (75% of erythrocytes), 2=a significant number of erythrocytes were hemolyzed (50% of erythrocytes), 3=a negligible number of erythrocytes were hemolyzed (25% of erythrocytes), and 4=complete fixation (absence of hemolysis). The CFT was considered positive when the score was 3 or 4.
Replication in cell lines
Replication of the Tr35 and Tr19 strains in various cell lines was tested using the Vero (African green monkey kidney cells), BHK-21 (Syrian hamster kidney fibroblast cells), and PEC (porcine embryonic cells) cell lines, which are most commonly used for arbovirus isolation. Cells were incubated at 37°C in a 5% CO2 atmosphere. Throughout the testing period, the cells were monitored at 24-h intervals to estimate the cytopathic effect (CPE), a sign associated with infection (Li et al. 2014).
Detection of a lipid envelope
The presence or absence of a lipid envelope was established by evaluating the median lethal dose (LD50/0.01mL) before and after ethyl ether treatment. Brain suspensions from infected mice with and without ethyl ether treatment were titered simultaneously with 10-fold dilutions, and the LD50/0.01mL was evaluated using the procedure described by Reed and Muench (1938). A decrease in the LD50/0.01mL value by several logarithms was the criterion for identifying susceptibility to ethyl ether treatment and, consequently, the presence of a lipid envelope (Klisenko 1986).
Electron microscopy
Brain suspensions were centrifuged at 4500 rpm for 30 min. The supernatants were pooled with an equal amount of homologous serum (diluted 1:10), incubated at 25°C for 2 h, and moved to 5°C for 18 h. After incubation, 1.5 mL of the mixture was centrifuged at 22,000 rpm at 10°C for 4 h. The supernatant was discarded, and the precipitate was dissolved in 150 μL of deionized RNase-free water (Qiagen).
Five microliters of this solution was absorbed onto a copper/carbon-coated grid (Mesh 400, Agar Scientific) for 1 h, stained with 2% ammonium molybdate (pH 5.1) for 20 s, rinsed in water, and dried prior to examination by electron microscopy using a JEM-100SX (Jeol) transmission electron microscope.
Nucleic acid extraction
Total nucleic acids were extracted and purified using the RIBO-prep DNA/RNA extraction kit (AmpliSens) according to the recommendations of the manufacturer. The DNA/RNA was eluted with 50 μL of RNase-free elution buffer (AmpliSens) and stored at −70°C.
Reverse transcription and amplification
Reverse transcription was performed using random hexanucleotide primers and the Reverta-L kit (AmpliSens) according to the manufacturer's recommendations. The cDNA was stored at −70°C and used as a template for amplification.
The full genomes of the Tr35 and Tr19 strains were sequenced (excluding the 5′ and 3′ ends of each segment). For this sequencing, a set of 26 primer pairs was used (Table 1). Primer design was based on the reference sequences for TRBV (HQ266581—HQ266590) (Dilcher et al. 2012). Overlapping fragments of the Tr35 and Tr19 genomes were amplified and sequenced. A hot-start amplification was performed in a 25-μL reaction containing 2 μL of cDNA, 1 μL of sense primer (10 pmol per μL), 1 μL of antisense primer (10 pmol/μL), 2.5 μL of dNTPs (1.76 mM, AmpliSens), and 10 μL of PCR buffer blue-2 (AmpliSens). The thermocycling parameters were set at 95°C for 2 min, followed by 40 cycles of 95°C for 15 s, 60–65°C for 20 s (depending on the primer pair), and 72°C for 30 s; a final elongation step was performed for 5 min. The reactions were performed in a MaxyGene gradient thermocycler (Axygen). The results of these amplification reactions were analyzed by 1.2% agarose gel electrophoresis containing ethidium bromide.
f, forward; r, reverse.
In silico analysis
Fragments were sequenced using the Sanger method on an ABI-Prism 3500 XL device (Applied Biosystems). Genomes were assembled using Lasergene 7.0 software (DNASTAR, Inc.). For the phylogenetic analysis, the VP1–VP10 sequences of the Tr35 and Tr19 strains were compared with sequences from appropriate strains of the Orbivirus genus. The GenBank accession numbers are provided in Table 2. The sequence alignments were performed using the CLUSTAL W software program (Thompson et al. 1994). The phylogenetic analysis was performed using the MEGA 6.0 software program with the Jukes–Cantor substitution model and the neighbor-joining (NJ) tree algorithm (Jukes and Cantor 1969).
Results
Virus identification
HA testing of the strains Tr35 and Tr19 resulted in a lack of hemagglutination with goose, sheep, and human type O erythrocytes over a pH range from 6.8 to 7.2. The results of the CFT for strain Tr35 are presented in Table 3. Acetone-extracted antigen from strain Tr35 was able to form immune complexes with antibodies against TRBV and KEMV (both viruses are members of the Orbivirus genus, Kemerovo serogroup). This resulted in strong complement-fixing activity, especially for the TRBV-antibody/strain-Tr35-antigen complex (1:64) and the strain-Tr35-antibody/Tr35-antigen complex (1:64). The complement-fixing activity for the KEMV-antibody/strain-Tr35-antigen complex was 1:32. The ability to form immune complexes with CNUV (a member of the genus Orbivirus, Chenuda serogroup) and CTFV (a member of the genus Coltivirus, Colorado tick fever serogroup) was not observed, and complexes were not observed with uninfected serum (complete lack of complement fixation). Therefore, strain Tr35 was identified as TRBV.
The results of the CFT for strain Tr19 are presented in Table 4. The complement-fixing activity for the TRBV-Tr35-antibody/Tr19-antigen complex was 1:8; the activity for the TRBV-Tr35-antibody/TRBV-Tr-35-antigen was 1:16. The complement-fixing activities of the TRBV-Tr19-antibody/TRBV-Tr-35-antigen and TRBV-Tr19-antibody/TRBV-Tr-19-antigen complexes were 1:64. The ability to form immune complexes with UUKV (a member of the genus Phlebovirus, family Bunyaviridae) and TBEV (a member of the genus Flavivirus, family Flaviviridae) was not observed, and complexes were not observed with serum from an uninfected mouse. Therefore, strain Tr19 was also identified as TRBV.
Replication in cell lines
A CPE was observed after 2 days of replication in the BHK-21 line and after 3 days in the PEC and Vero lines.
Detection of a lipid envelope
Comparative evaluations of the LD50/0.01mL for the strains Tr35 and Tr19 showed susceptibility to ethyl ether treatment. According to our data, the LD50/0.01mL for strain Tr35 was 6.2 logs before treatment and 5.0 logs after treatment, whereas the LD50/0.01mL for strain Tr19 was 7.0 logs before treatment and 5.2 logs after treatment. Therefore, there was a 1.2-log reduction in the LD50/0.01mL for strain Tr35 and a 1.8-log reduction in the LD50/0.01mL for strain Tr19. These data confirmed the absence of a lipid envelope in both strains.
Electron microscopy
The virus particles were approximately 70 μm with a defined surface structure containing ring-shaped capsomeres (see Fig. 2). This structure is consistent with viruses of the genus Orbivirus within the family Reoviridae (Mertens et al. 2000).

Electron micrograph of virus particles. The size of particles is approximately 70 μm.
Phylogenetic analysis
The phylogenetic analysis of the VP1–VP10 nucleotide sequences of the Tr35 and Tr19 strains, along with the appropriate sequences of other orbiviruses, confirmed their classification in the genus Orbivirus (Figs. 3 and 4). The location of Tr35 and Tr19 on the phylogenetic trees indicates their membership in the GIV subgroup and a close relationship with the TRBV that was sequenced previously (Dilcher et al. 2012), as well as with LIPV and KEMV. The pairwise comparison of the VP1 sequences within the GIV subgroup demonstrated that Tr35 and Tr19 share 89% nucleotide identity with each other. Strain Tr35 shares 94% identity with the reference strain TRBV, whereas strain Tr19 shares 90% identity with the reference strain. Strain Tr35 shared 94% identity with LIPV, whereas strain Tr19 was 89% identical to LIPV. For both the Tr35 and Tr19 strains, the shared identities with the reference strain KEMV, strain KEMV-21/10 and GIV were 70%, 71% and 68%, respectively (see Table 5).

Phylogenetic tree of the Orbivirus genus based on VP1 gene sequences (nucleotide level). The phylogenetic analysis was performed using the Jukes–Cantor substitution model, and the trees were reconstructed using the neighbor-joining (NJ) method in the MEGA 6.0 software. The statistical significance of the tree topology was evaluated by bootstrap resampling of the sequences 1000 times. The locations of the Tr35 and Tr19 strains are marked with black dots on the tree. BTV, bluetongue virus; EHDV, epizootic hemorrhagic disease virus; PATAV, Pata virus; TIBOV, Tibet orbivirus; EUBV, Eubenangee virus; TILV, Tilligerry virus; CGLV, Changuinola virus; LEBV, Lebombo virus; EEV, equine encephalosis virus; ORUV, Orungo virus; KASV, Kasba virus; AHSV, African horse sickness virus; YUOV, Yunnan virus; MPOV, Mobuck virus; PHSV, Peruvian horse sickness virus; SVIV, Sathuvachari virus; CORV, Corriparta virus; UMAV, Umatilla virus; SLOV, Stretch Lagoon orbivirus; OKHV, Okhotskiy virus; GIV, Great Island virus; KEMV, Kemerovo virus; TRBV, Tribeč virus; LIPV, Lipovnik virus.
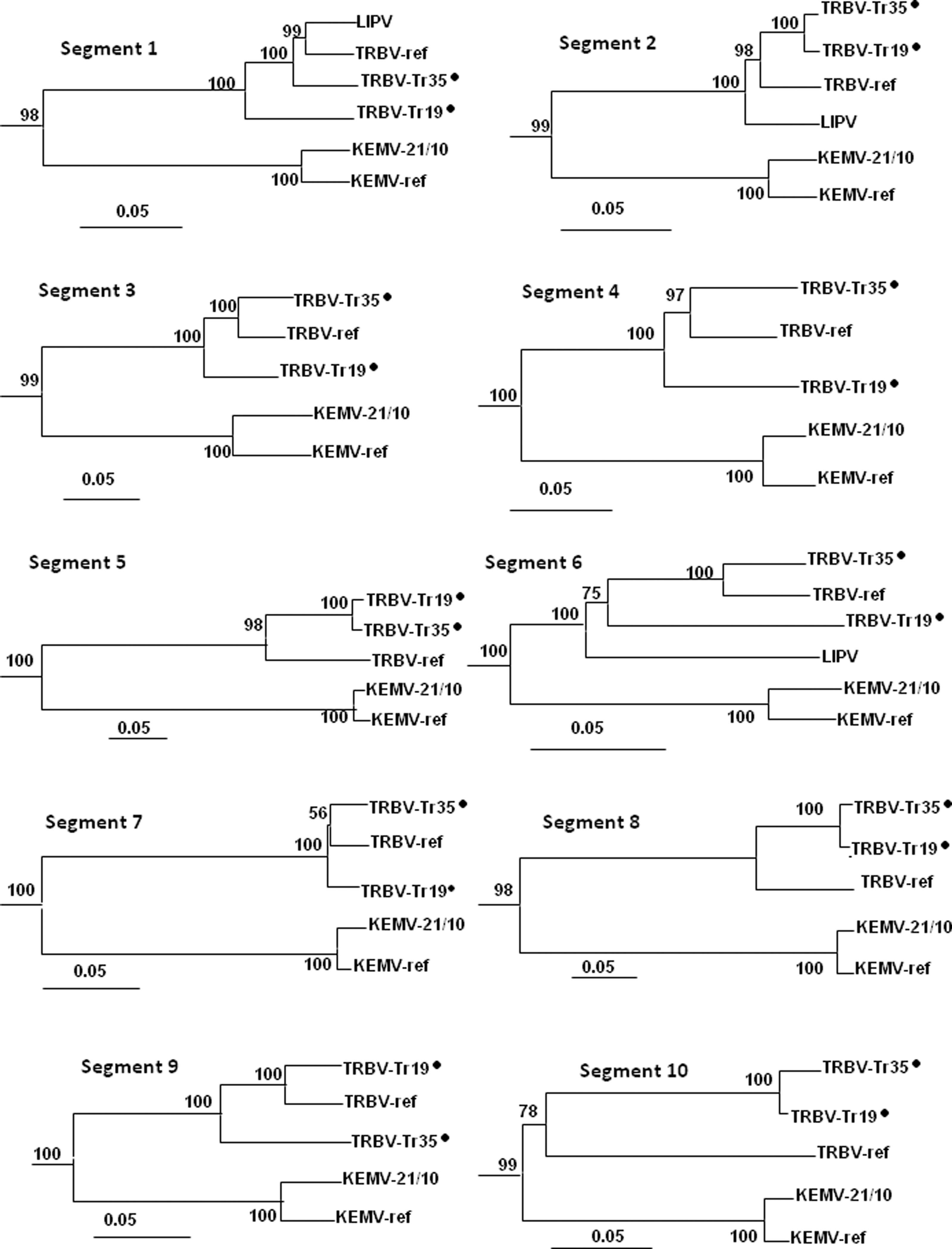
Phylogenetic tree of the Great Island virus (GIV) subgroup based on VP1–VP10 gene sequences (nucleotide level). The phylogenetic analysis was performed using the Jukes–Cantor substitution model, and trees were reconstructed using the neighbor-joining (NJ) method and MEGA 6.0 software. The trees were rooted using the appropriate sequences of the GIV segments, and the GIV branches were deleted from the trees. The statistical significance of the tree topology was evaluated by bootstrap resampling of the sequences 1000 times. The locations of the Tr35 and Tr19 strains are marked with black dots on the trees. LIPV, Lipovnik virus; TRBV, Tribeč virus; KEMV, Kemerovo virus.
TRBV, Tribeč virus; LIPV, Lipovnik virus; KEMV, Kemerovo virus; GIV, Great Island virus.
Comparisons of the VP3 sequences demonstrated a 99% nucleotide identity between strains Tr35 and Tr19 and a 95% identity between the Tr35 and Tr19 strains and the reference sequence of TRBV. The shared identities with the sequences of LIPV, KEMV, and GIV were 93%, 76%, and 73%, respectively.
Phylogenetic trees for different genomic segments revealed distinct phylogenetic relations among TRBV-ref, Tr19, and Tr35 strains. In segments 1, 3, 4, 6, and 7, the Tr19 strain was an outgroup to the reliably supported group of TRBV-ref and Tr35 strains. In segments 2, 5, 8, and 10, TRBV-ref was an outgroup to the reliably supported group of Tr19 and Tr35. In segment 9, the Tr35 strain was an outgroup to TRBV-ref and Tr19 strains. Moreover, different phylogenetic positions of the LIPV strain sequences were observed within the GIV subgroup. LIPV was an outgroup to the three TRBV sequences in segments 2 and 6, but grouped reliably with TRBV in segment 1. Unfortunately, we could not extend this observation to other genome segments, because there was only sequence of three genome regions available in GenBank.
Discussion
Here we report the isolation and characterization of two strains of TRBV isolated in Ukraine in 1988 and 2008. The affiliation of both strains with the genus Orbivirus within the Reoviridae family was confirmed using CFT, electronic microscopy, and molecular methods. In addition, CFT demonstrated the presence of cross-reactivity with ascites fluid against KEMV, a virus closely related to TRBV, and the absence of cross-reactivity with ascites fluid against CNUV and CTFV, as well as with sera against TBEV and UUKV. The electron microscopy study demonstrated viral particles with shapes and sizes distinctive for orbiviruses.
HA testing of strains Tr35 and Tr19 showed a lack of hemagglutination with goose, sheep, and human type O erythrocytes. Thus, HA testing could not be used for diagnostic testing of TRBV (Hubálek and Rudolf 2012). Susceptibility to treatment with ethyl ether confirmed the absence of a lipid envelope, which is typical for Reoviridae (Mertens et al. 2000). The two strains demonstrated the ability to replicate in BHK-21, PEC, and Vero cell lines. During replication in the BHK-21 line, a CPE was observed slightly earlier than in the PEC and Vero lines.
Full genomes of the Tr35 and Tr19 strains were sequenced (excluding the 5′ and 3′ ends of each segment) and submitted to GenBank under the accession numbers KJ574044–KJ574045 and KJ010789–KJ010806. The phylogenetic analysis demonstrated the close relationships of Tr35 and Tr19 with the reference TRBV strain. According to the Ninth Report of the International Committee on Taxonomy of Viruses, a species demarcation criterion in the genus Orbivirus is less than 74% identity among the VP3 nucleotide sequences (King et al. 2012). An analysis of the sequence identities for TRBV, KEMV-ref, KEMV-21/10, LIPV, strain Tr35, and strain Tr19 demonstrated that the identities ranged from 76% to 95% (Table 5). Therefore, all of these strains should be considered members of one species according to current criteria. Phylogenetic analysis revealed distinct and reliably supported grouping patterns among TRBV and LIPV in different genome segments. This indicates that segment reassortment was common in TRBV, and supports assignment of LIPV to TRBV, because their genome segments were genetically compatible and could be involved in reassortment. No phylogenetic evidence of reassortment between TRBV and KEMV was observed. The sample of five genomic sequences does not allow such reassortment to be completely ruled out, but if future studies do not reveal reassortment between KEMV and TRBV, this would speak for acknowledging them as distinct taxonomic entities.
For a more detailed understanding of the complicated phylogeny within the GIV subgroup, epidemiological studies are needed, and additional isolates of TRBV and KEMV should be analyzed.
Footnotes
Acknowledgments
This work was supported by the Russian Federal Target Program “National system of chemical and biological safety of the Russian Federation (2009–2013)” (project no. 60-D 17072012). The authors express deep gratitude to D.K. Lvov, the head of the D.I. Ivanovsky Institute of Virology, for his assistance in confirming the identification of strain Tr35.
Author Disclosure Statement
No competing financial interests exist.