
Abstract
Arenavirus Sabiá was originally isolated from a fatal human infection in Brazil, and after the occurrence of the second fatal human case in São Paulo state, epidemiologic and virologic studies were performed in the area where the patient lived, aiming at the identification of the Sabiá natural rodent reservoir. A broadly cross-reactive enzyme-linked immunosorbent assay (ELISA) was used to screen for antibody-positive samples. Antibodies to arenavirus were detected in two of the 55 samples of Calomys tener, and from these results, samples of rodents were analyzed by a broad RT-PCR assay. RT-PCR amplification detected arenavirus sequences in five of the 55 C. tener samples, and sequencing showed that this virus is a distinct form of Sabiá virus. Thus, we describe here the evidence for the circulation of a new arenavirus in Brazil (proposed name Pinhal virus) and its genetic characterization compared to other arenaviruses. This study also suggests C. tener as a probable rodent reservoir for this virus and associates this new virus with the lineage C of New World arenaviruses. Although we have defined some characteristics of this virus, so far, there is no evidence of its involvement in human disease.
Introduction
T
Arenaviruses have ambisense single-stranded RNA genomes consisting of two segments known as large (L) and small (S). Phylogenetic analysis of nucleotide sequences of the S segment indicates that the New World arenaviruses are classified into three lineages, designated A, B, and C. (Bowen et al. 1996, Archer et al. 2002, Gonzalez et al. 2007). Some of the viruses belonging to lineages A and B are pathogenic to humans, and some lineage B viruses cause viral hemorrhagic fever; however, no virus from lineage C has been associated with human disease. The arenaviruses known to occur in Brazil are: Flexal (FLEV) (Pinheiro et al. 1977), Amapari (AMAV) (Pinheiro et al. 1966), and Cupixi (CPXV), all associated with Oryzomys capito, and SABV (Coimbra et al. 1994), whose rodent host is unknown to date.
SABV was initially reported from São Paulo State and since then has been isolated only from two fatal hemorrhagic cases and another two laboratory-acquired infections in persons working directly with the virus (Vasconcelos et al. 1993, Barry et al. 1995). The first reported fatal human case was a 25-year-old woman from Cotia County, in 1990 (Coimbra et al. 1994); the second case was a 32-year-old-man from Espírito Santo do Pinhal County, in 1999 (Coimbra et al. 2001). Both patients reported contact with rural environments, but the regions where the patients lived are about 200 km apart, showing that SABV may be disseminated throughout São Paulo State. Following the investigation of the second fatal case associated with SABV, epidemiological and virological studies were established in the area where the patient lived, aiming at the identification of potential rodent reservoirs for SABV in São Paulo State and evidence for its circulation in the region.
Captures of rodents were performed in Espirito Santo do Pinhal County as part of a research project for molecular characterization and phylogenetic analysis of hantavirus and arenavirus occurrence in that area, characterized as a remainder of the Brazilian Atlantic Forest ecosystem. Among 1298 blood samples and tissues of rodents captured for the project, 412 were from the municipality of Espirito Santo do Pinhal. As a result of this work, a putative new arenavirus was isolated from that area, and this virus might have been circulating in the area for quite some time.
Material and Methods
Ethics and biosafety
Field activities concerning rodent trapping and handling were cleared by national legal provisions. The collection and handling of biological samples were carried out following the recommendations of Mills et al. (1995), applying humane methods of anesthesia and euthanasia. The research project was approved by National Council for Technological and Scientific Development–CNPq, which fully considered the ethical aspects in granting the research funds.
Study site and animal trapping
Rodent trapping was conducted in the natural habitats located in Espírito Santo do Pinhal County (22°11′27″S 46°44′27″W), on the west slope of the Serra da Mantiqueira, around 200 km north from São Paulo City (Fig. 1). Rodents were trapped by using Sherman live traps in two time points—May of 2005 and February of 2006. Blood and tissue samples, including lung, kidney, liver, spleen, heart, and brain, were obtained from each rodent. Carcasses were tentatively identified in the field and preserved in a 10% formalin solution for further taxonomic confirmation.
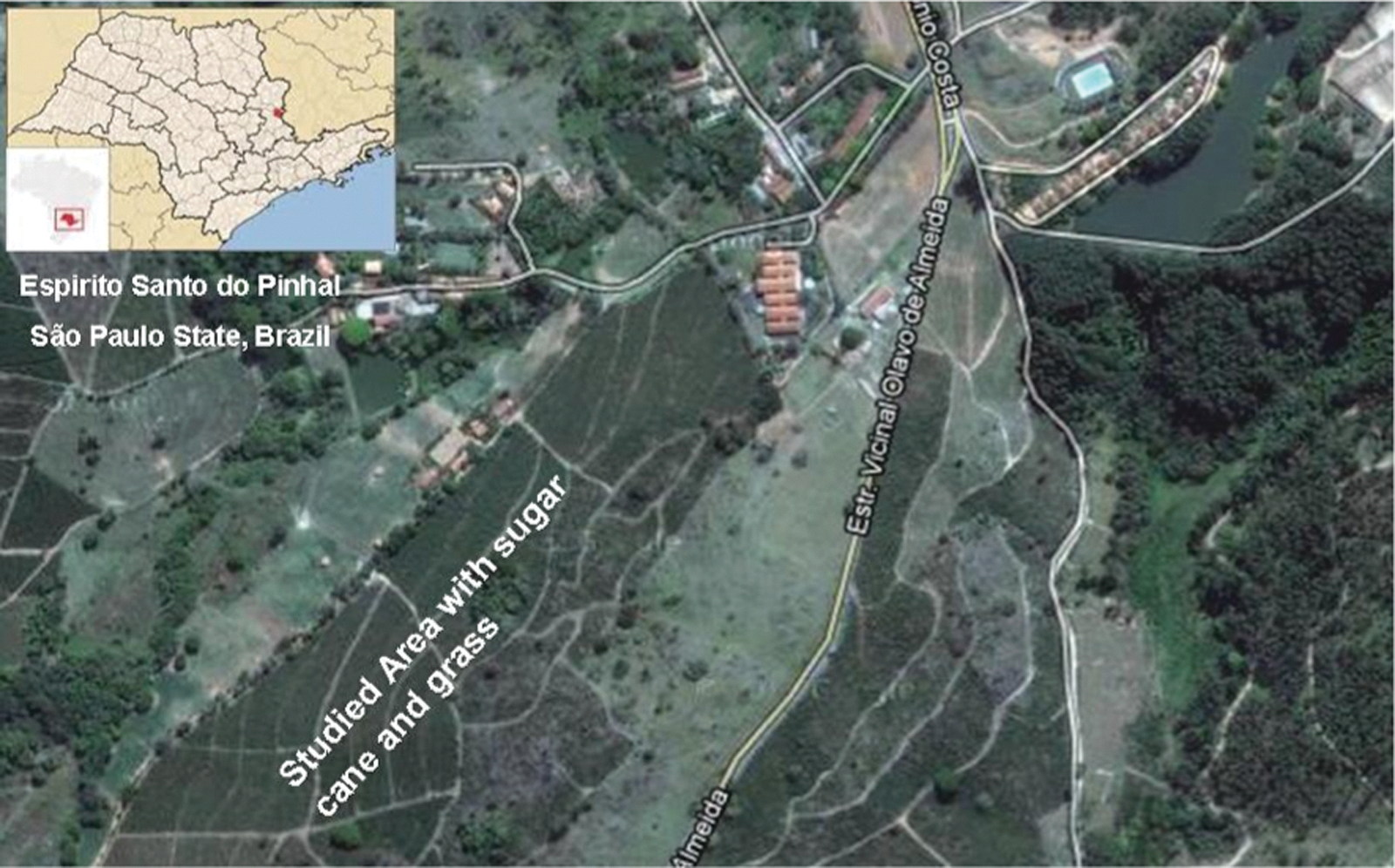
Location of study area in São Paulo State, Brazil. Color images available online at
Serologic investigation
Blood samples from captured rodents were analyzed by a broad-reactive arenavirus enzyme-linked immunosorbent assay (ELISA) to detect immunoglobulin G (IgG) antibodies against arenaviruses (Morales et al. 2002). Arenavirus antigen, prepared using detergent extraction from Vero E6 cells infected with the XJCl3 JUNV strain, was provided by the Instituto Nacional de Enfermedades Virales Humanas Dr. Julio I. Maiztegui, Pergamino, Argentina. A pool of peroxidase-labeled secondary antibodies (anti–Rattus norvegicus, anti–Peromyscus leucopus, anti-Mouse) and the ABTS Peroxidase Substrate System (contains 2,2′-azino-di-[3-ethylbenzthiazoline-6-sulfonate] and hydrogen peroxide [H2O2]) as a substrate were used in the ELISAs. The reading of optical density was made in an ELISA reader (Bio-Rad), using a 405-nm filter.
The rodent samples were processed at a dilution of 1/400. The sample was considered positive when the value of optical density was greater than the cutoff value for the test. Cutoff values were determined as the mean value of five readings using uninfected rodent sera plus three times the standard deviation (SD) of each test. IgG detection by this ELISA was the screening methodology used to investigate probable rodent reservoirs species to arenaviruses.
Molecular investigation for arenavirus infection in rodent tissue samples
Total RNA was extracted from rodent brains, individually or in pool of five or three animals, using the QIAamp RNA Blood Kit (QIAGEN Inc., Ontario, Canada), following the manufacturer's instructions.
Viral RNA was detected by one-step RT-PCR using oligonucleotides that amplify a fragment of 580 nucleotides of the nucleoprotein (NP) gene located on the S segment, by using generic primers as described by Bowen et al. (1996). The PCR products of the expected size were separated on 1.5% agarose gel electrophoresis and stained with ethidium bromide. Both strands of the purified PCR products were directly sequenced in an ABI 377 automated sequencer using the BigDye terminator sequencing version 3.1 kit (Applied Biosystems, Foster City, CA). The nucleotide sequences from this study were deposited in the GenBank under the accession numbers: EU220740 (SPAn 266400), EU280545 (SPAn 266403), EU280546 (SPAn 266468), and EU2805547 (SPAn 266474).
Due reasons of safety, there were no attempts to culture virus.
Phylogenetic analyses
DNA sequences were edited using both Chromas version 1.45 and EditSeq version 5.00 software (Lasergene DNAStar Inc. package), and alignment of nucleotide sequences was performed with the BioEdit program by the Clustal Wallis method (
The DNA fragments were aligned with representative sequences of the arenavirus genotypes retrieved from GenBank (Table 1). Bayesian phylogenetic reconstructions were performed using Markov Chain Monte Carlo analysis implemented by BEAST version 1.6.2 (Drummond and Rambaut 2007). The internal nodes were inferred using a Markov Chain Monte Carlo (MCMC) Bayesian approach under a general time reversible (GTR) model with Gamma-distributed rate variation (γ) and a proportion of invariable sites (I), using a relaxed (uncorrelated lognormal) molecular clock. Four independent MCMC runs of four chains each were ran for 10 million generations. We used a Bayesian Skyline prior to estimating population dynamics through time (Drummond et al. 2005) and accessed an estimative of evolutionary rate and the time to the most recent common ancestor (TMRCA) (Drummond et al. 2006).
Results
Among 1298 blood samples and tissues of rodents captured for the project, 412 were from the municipality of Espirito Santo do Pinhal. Antibodies to hantavirus were detected in 46 rodent samples. Excluding samples positive for hantavirus, 1252 samples were processed for the detection of antibodies and amplification of the arenavirus genome. Arenavirus antibodies were only found in two of the 55 serum samples of C. tener. A total of 366 pools were made containing three or five fragments of brain and spleen, organized by species and sample origin, and each pool of the 366 organ pools was tested separately. Suffice to say that different tissues of each rodent captured were collected, but the RT-PCR for arenaviruses were performed only on fragments of brain and spleen. Other tissues collected were used for hantavirus research.
PCR amplification detected arenavirus sequences in five (9%) of the 55 Calomys tener samples, and amplicon sequencing provided evidence for a new arenavirus, denominated hereafter Pinhal virus (PINV). Table 2 shows nucleotide and amino acid identities of PINV strains compared to other arenaviruses, either from Brazil or from other lineage C viruses, and with prototype LCMV. PINV strains are distinct from other arenaviruses of lineage C (average difference, 27.5% [nucleotides]) and 20.5% [amino acids]), and from other Brazilian arenaviruses (39.7% [nucleotides] and 39.9% [amino acids]). The difference between PINV and other lineage C arenavirus (Latino and Oliveros, for instance) are sufficient to reinforce the view that PINV has a distinct genetic identity value to be considered a separate virus from Latino and Oliveros viruses.
Upper right=nucleotide distances (%); lower left=deduced amino acid distances (%).
MACV, Machupo virus; SABV, Sabiá virus; AMAV, Amapari virus; FLEV, Flexal virus; CUPV, Cupixi virus; PARV, Parana virus; JUNV, Junin virus; OLVV, Oliveros virus; PAMV, Pampa virus; LATV, Latino virus; GUAV, Guanarito virus; PIRV, Pirital virus; PICV, Pichinde virus; ALLV, Allpahuayo virus; TACV, Tacaribe virus; WWAV, Whitewater Arroyo virus; TAMV, Tamiami virus; BCNV, Bear Canyon virus; LCM, lymphocytic choriomeningitis virus; LASV, Lassa virus.
The phylogenetic analysis recovered a strongly supported tree, with a general posterior probability of 0.89 supported by an effective sample size (ESS) of 482 after four MCMC runs, where all ESS parameters were above 150. Figure 2 shows the MCMC consensus tree obtained for the New World arenaviruses. Our data show that PINV strains constitute a subclade within a lineage C clade (posterior probability of 0.998%) and support the monophyletic relationship of these viruses, indicating that PINV is associated with the arenavirus branch that is nonpathogenic to humans.
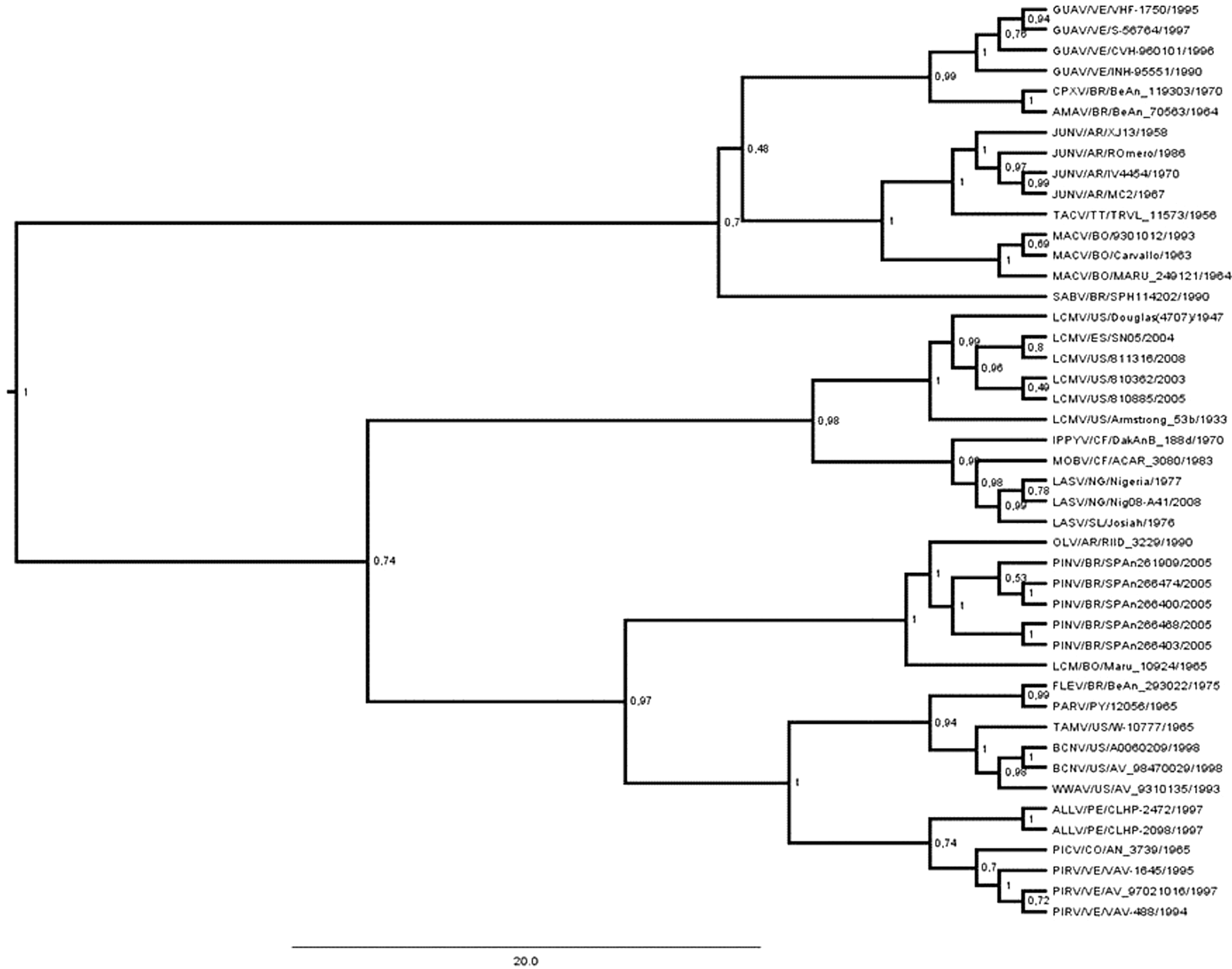
Markov Chain Monte Carlo (MCMC) consensus tree obtained for the New World arenaviruses supported by an effective sample size (ESS) of 482 after four MCMC runs, where all ESS parameters are above 150. Phylogenetic analysis recovered a strongly supported tree (general posterior probability of 0.89).
Molecular clock analysis suggests that PINV emerged in the last 100 years, with TMRCA calculated around 121 (SD=12 years) years ago. The substitution rate for arenavirus evolution was estimated to be around 1.567*10−3 nucleotides/site/year (SD=7.5181*10−5), an expected result for a RNA virus.
Discussion
Arenaviruses are emerging viruses that are associated with some severe diseases, and their transmission to humans is related to inhalation of aerosolized rodent excreta. However, for some of these viruses, such as SABV, their rodent hosts are unknown. This article describes a newly detected arenavirus and its putative rodent reservoir during a field research to elucidate the SABV reservoir. The proposed name for this virus is PINV, based on the location where the rodents were captured.
Rodents were captured during an investigation of a fatal human case associated to SABV infection. Among the 55 individuals of the species C. tener captured in the county of Espírito Santo do Pinhal, two had cross-reactive antibodies to JUNV by ELISA. In five other specimens caught near the capture sites of ELISA-positive rodents, it was possible to amplify the PINV genome. All seven positive rodents were captured during the 2005 expeditions. These results might be evidence that this species may act as rodent reservoir for arenaviruses in this region. Another interesting finding was the discordance of the ELISA and the RT-PCR results, because PINV was not detected in rodents positive for arenavirus antibodies. The reason for these results is not clear, but we can assume that our RT-PCR is not sensitive enough to detect virus in rodents with high titers of circulating antibodies, because these antibodies might reduce the viral load on the rodents. Alternatively, one can assume that there is another arenavirus circulating in that region and it is more closely related to JUNV than PINV, because the antigen used on the ELISA investigation was constituted by JUNV extracts.
The rodents captured on this study represent a sample from a mixed community of species from an extensively modified habitat (Table 3). Species composition reflects the influence of fragmentation and anthropization of the environment with a low diversity of rodents and a marked dominance of Necromys and Akodon. The presence of Mus musculus and Rattus rattus is a clear indication of the influence of human populations in this environment. In this context, the detection of an arenavirus in close proximity to human dwellings poses an increased risk of transmission of rodent-borne diseases, and protective measures should be reinforced in these areas. Although these changes in environment create the grounds for transmission of rodent-borne viruses, and the arenavirus described here was detected during an investigation of a fatal infection by SABV, phylogenetic analysis of the nucleotide sequence indicates that PINV groups with the nonpathogenic Oliveros and Latino viruses within the lineage C in the New World arenavirus complex (Bowen et al. 1996). Because this might indicate that PINV is not associated with human disease, further serological surveys and increased surveillance are needed to determine whether or not PINV can infect humans. Thus, epidemiological and virological studies on the localities where C. tener and PINV coexist, including the use of molecular techniques, must be carried out in the future. Furthermore, the phylogenetic analysis based on the NP nucleotide sequence data indicate that SPAn 266400, SPAn 266403, SPAn 266468, and SPAn 266474 are PINV strains associated with C. tener in São Paulo State.
C. tener is a small rodent adapted to “cerrado,” a characteristic savannah-like ecosystem common to the central region of Brazil. These rodent populations are highly favored by the extensive sugar cane culture and by the growing of pasture. In our study, it was the fourth most frequently captured species, accounting for 13.3% of the sampled rodents.
Despite the fact that the Calomys genus is widely spread in this country, it is not known yet which virus shares the same geographic distribution. On the basis of the known geographic distribution of other arenaviruses and by the lack of evidence of its circulation in other locations studied during this investigation, it is possible that Pinhal arenavirus may be restricted only to a portion of its natural reservoir territory. This assumption is corroborated by the fact that arenaviruses have a limited geographic distribution, as it is the case for most of the New World arenaviruses. The occupancy of a restricted area within their natural reservoir distribution, known as natural nidality (Salazar Bravo et al. 2002), may have helped the detection of PINV, and may represent the most significant barrier to uncover the SABV reservoir puzzle. However, because there is evidence that SABV circulates in the same geographic region as PINV, as it was recently isolated from a hemorrhagic human case from Espírito Santo do Pinhal county (Coimbra et al. 2001), other cases of human disease potentially attributed to an arenavirus infection in this region must be investigated carefully. However, some obstacles must be overcome during this investigation because there is a considerable cross-reactivity among New World arenaviruses in ELISA tests, such as the one observed here, e.g., JUNV with SABV and PINV. Thus, in the search for an etiologic agent of a hemorrhagic fever, the investigation of new cases in the region must include additional diagnostic methods, such as viral isolation, neutralizing antibodies detection, and viral nucleic acid amplification.
Further studies on arenavirus distribution in Brazil are needed, initially to determine how much PINV is antigenically distinct from SABV and to define their detailed phylogenetic and phylogeographic relationship. It is critical to describe the real geographical distribution of SABV because the two described fatal cases lived in distinct regions in São Paulo State and for this reason it is necessary identify the species reservoir. The patient who lived in Cotia County reported extensive traveling, but the second case was a resident of Espírito Santo do Pinhal who had not traveled outside the region.
Because both SABV cases occurred many years apart from each other, it might be either an incidental nature of the transmission or expansion of SABV circulation. Other arenaviruses are associated with epidemic transmission, mostly due to the intrinsic association of rodent reservoirs with agricultural activities. This was true regarding GTOV and the cases associated with it during the outbreak that occurred in Portuguesa State in Venezuela (Tesh et al. 1994), as well as the transmission of JUNV in Argentina and MACV in Bolivia.
Elucidating the natural geographic distribution of PINV and SABV is a matter of concern to the Brazilian public health system, because other arenaviruses might be associated with serious epidemics in relatively restricted environments and little is known to positively label PINV or even SABV as unassociated with outbreaks or clustered cases. The detection of these two viruses, even infrequently, might be understood as a marker of the intensification of rodent/human interactions, probably leading to transmission of arenaviruses in Brazil.
Footnotes
Acknowledgments
This study was partially supported by funds from CNPq/Proc. 403023/2004-1 and São Paulo State Health Secretary.
The authors would like to thank the staffs of Núcleo de Doenças de Transmissão Vetorial of Instituto Adolfo Lutz, São Paulo/SP, Brazil, Research Department of the Instituto Nacional de Enfermedades Virales Humanas Dr. Julio I. Maiztegui, Pergamino, Argentina, and Centro de Controle de Zoonoses–Espírito Santo do Pinhal County, whose work made this study possible.
Author Disclosure Statement
The authors declare that no financial or other competing interests exist.