
Abstract
Multidrug-resistant bacteria are a growing problem in different environments and hosts, but scarce information exists about their prevalence in reptiles. The aim of this study was to analyze the resistance mechanisms, molecular typing, and plasmid content of cefotaxime-resistant (CTXR) Escherichia coli isolates recovered from cloacal samples of 71 turtles sheltered in a herpetarium in Mexico. CTXR-E. coli were recovered in 11 of 71 samples (15.5%), and one isolate/sample was characterized. Extended-spectrum β-lactamase (ESBL)-producing E. coli isolates were detected in four samples (5.6%): two strains carried the bla CTX-M-2 gene (phylogroup D and ST2732) and two contained the bla CTX-M-15 gene (phylogroup B1 and lineages ST58 and ST156). The bla CMY-2 gene was detected by PCR in E. coli isolates of eight samples (9.8%) (one of them also carried bla CTX-M-2); these isolates were distributed into phylogroups A (n = 1), B1 (n = 6), and D (n = 1) and typed as ST155, ST156, ST2329, and ST2732. Plasmid-mediated quinolone resistance (PMQR) genes were detected in five isolates [aac(6′)Ib-cr, qnrA, qnrB19, and oqxB]. From three to five replicon plasmids were detected among the strains, being IncFIB, IncI1, IncFrep, and IncK the most prevalent. ESBL or pAmpC genes were transferred by conjugation in four strains, and the bla CTX-M-15 and bla CMY-2 genes were localized in IncFIB or IncI1 plasmids by Southern blot hybridization assays. Class 1 and/or class 2 integrons were detected in eight strains with six different structures of gene cassette arrays. Nine pulsed-field gel electrophoresis patterns were found among the 11 studied strains. To our knowledge, this is the first detection of ESBL, CMY-2, PMQR, and mobile determinants of antimicrobial resistance in E. coli of turtle origin, highlighting the potential dissemination of multidrug-resistant bacteria from these animals to other environments and hosts, including humans.
Introduction
M
Recently, it has been suggested that antibiotic-resistant bacteria isolated from aquatic animals, as Chelonia mydas and Caretta caretta sea turtles species, can be used as bioindicators for pollution in the coastal marine habitats, which is considered to be an ecological problem (Foti et al. 2009, Al-Bahry et al. 2012). However, to our knowledge, there does not exist information about the presence of ESBL-, pAmpC-, or PMQR-producing E. coli isolated from turtles, or about the potential role of these animals as a source of multidrug resistance bacteria in the household environment. For this reason, and due to the increasing popularity and variety of turtles as pets in recent years, the aim of this study was to identify antimicrobial resistance genes, characterization of integrons and plasmids, as well as the molecular typing of cefotaxime-resistant (CTXR) E. coli isolated from sheltered turtles in Puebla, Mexico.
Materials and Methods
Animals tested
Cloacal swabs were taken in 2012 from all 71 healthy adult turtles sheltered in the herpetarium of the Benemérita Universidad Autónoma of Puebla (Puebla, Mexico). There were 34 female and 37 male turtles of Kinosternon integrum (n = 37), Trachemys scripta elegans (n = 30), Chelydra serpentine (n = 3), and Rhinoclemmys spp. (n = 1) species. The turtles came from environmental habitats (n = 4) collected by students during camping practices or from household environments, in which turtles were considered as pets and later were donated by their owners (n = 67). In the herpetarium, they were fed with chicken liver, vegetables, mealworms, and commercial turtle biscuits. The cloacal samples (one per turtle) were transported in Stuart medium to the laboratory and were maintained in refrigeration at 4°C until processing, within a 48-h period.
Isolation of E. coli strains
The samples were inoculated on Levine agar plates supplemented with cefotaxime (2 μg/mL) for CTXR E. coli recovery. CTXR bacteria with features corresponding to E. coli were randomly selected (up to three per sample) and identified by standard biochemical tests (Gram staining, lactose fermentation, indol production, and methyl red/Voges–Proskauer test) and by the species-specific PCR for uidA (beta-glucuronidase enzyme) gene detection (Ben Sallem et al. 2014).
Antimicrobial susceptibility testing
Susceptibility testing was performed by the disk diffusion method, and the susceptibility was interpreted according to the Clinical Laboratory Standards Institute (CLSI, 2013), for ampicillin, amoxicillin/clavulanate, cefalotin, cefoxitin, cefotaxime, ceftriaxone, ceftazidime, cefepime, aztreonam, imipenem, meropenem, nalidixic acid, ciprofloxacin, gentamicin, amikacin, tobramycin, streptomycin, chloramphenicol, trimethoprim, trimethoprim/sulfamethoxazole, and tetracycline. E. coli ATCC 25922 was used as a control strain.
The screening of phenotypic ESBL production was performed with the standard double-disk synergy test, using disks of cefotaxime (30 μg), ceftazidime (30 μg), aztreonam (30 μg), and cefepime (30 μg), with a centrally positioned disk of amoxicillin/clavulanate (20/10 μg) at a center-to-center distance of 25–30 mm. The presence of clear-cut extension of the edge of β-lactamic inhibition zone toward the disk containing β-lactamase inhibitor was considered as a positive test for ESBL.
Characterization of antimicrobial resistance genes and integrons
β-lactamase (bla CTX-M groups 1, 2, 8, and 9, bla OXA, bla SHV, and bla TEM) and PMQR encoding genes [qnrA, qnrB, aac(6′)-Ib-cr, qepA, oqxA, and oqxB], as well as aminoglycoside [aac(3)-II, aadA1/2], sulfonamide (sul1, sul3), chloramphenicol (cmlA), trimethoprim (drfA), and tetracycline [tet(A), tet(B)] resistance genes were tested by PCR and sequencing in all the recovered CTXR E. coli isolates (Vinué et al. 2009, Ruiz et al. 2012).
The presence of plasmid-mediated AmpC β-lactamase genes was tested by using a multiplex PCR (Pérez-Pérez and Hanson 2002). Mutations in the regulatory region of ampC chromosomal gene were determined by PCR and sequencing, as well as the mutations in the gyrA and parC genes in the quinolone-resistant isolates (Ruiz et al. 2012). The presence of intI1 and intI2 genes was tested by PCR and their variable region structures were determined by PCR and sequencing following the primer walking strategy (Sáenz et al. 2010).
Phylogenetic grouping
E. coli isolates were classified in the phylogenetic groups according to the Clermont phylotyping method, which is based on a PCR assay to detect the genes chuA, yjaA, arpA, and the DNA fragment TspE4.C2 (Clermont et al. 2013).
Multilocus sequence typing and pulsed-field gel electrophoresis typing
The molecular typing of selected strains was carried out by multilocus sequence typing (MLST) analysis, sequencing internal fragments of seven standard housekeeping genes (adk, fumC, icd, purA, gyrB, recA, and mdh), and getting the allele combination at
All the strains were also typed by pulsed-field gel electrophoresis (PFGE) with XbaI (Vinué et al. 2008), PFGE patterns were classified by the cluster analysis of Dice similarity index (value >85%) to assign a pulsotype (PT), and a dendogram was also generated by Dice/Unweighted Pair Group Method using arithmetic averages (UPGMA) analysis using the NTSYSpc 2.21q software.
Conjugation assays
Conjugation experiments were carried out to determine the transferability of ESBL and pAmpC encoding genes in nine selected donor strains (based on PFGE patterns and genotypic resistance determinants), using the rifampicin-resistant (RIFR) E. coli C600, RIFR E. coli J53, or azide-resistant (AZR) E. coli J53 as recipient strains. Transconjugants were selected in MacConkey agar plates supplemented with rifampicin (100 μg/mL) or azide (100 μg/mL) and cefotaxime (2.5 μg/mL). Transconjugants were tested for resistance to antimicrobial agents, phenotypic ESBL production, and PCR for detection of ESBL, pAmpC, and other resistance genes, as described above.
Plasmid characterization
Plasmids were classified according to their incompatibility group using the PCR-based replicon typing method, which comprises the major plasmid incompatibility groups among Enterobacteriaceae (Carattoli et al. 2005). PFGE with S1 nuclease (S1 Nuclease Fermentas Thermo Scientific) digestion of whole genomic DNA was performed for all 11 donor strains and transconjugants to determine the plasmid content and size (Ben Sallem et al. 2014), compared with MidRange I PFG Marker (New England Biolabs). The plasmid-containing reference strain NCTC 50192 was used as a control strain (Philippon et al. 1997), and the plasmid sizes (PSs) were estimated using statistical analysis. A simple linear regression analysis through the least squares method was performed to determine the PSs, using the Statgraphics software (Statgraphics 2010) for data processing. This software designed a calculation model that approximated the expected PS as a function of the experimental data of migration distance, compared with the size marker (Midrange PFG Marker) migration in the S1-PFGE gel. The resulting linear regression equation was PS = (23.1383 − 0.245662 × DM)2 with r 2 = 99.96, which explains the variability of molecular size as a function of the migration distance with a 95% confidence interval (absolute error = 0.05).
Hybridization analysis
To test the plasmid localization of transferred resistance determinants, hybridization experiments with radioactive labeling were carried out in plasmid DNA of donor and transconjugant strains. Southern blots were performed from resolved plasmids obtained by the S1-PFGE procedure, which were transferred to nylon membranes (Hybond-N; GE Healthcare Lifescience) with a 0.4 M NaOH solution. Prehybridizations were assayed at 42°C for 4 h with the prehybridization solution (Formamide, 20 × SSC, 100 × Denhards, 0.25 M EDTA pH8, 1.0 M Tris pH 7.5, sodium dodecyl sulfate [SDS] 10%, calf thymus DNA, H2O). Hybridization using 32P labeling and detection kit (RadPrime DNA Labeling System; Thermo Fisher Scientific) was performed with PCR-generated probes specific for bla
CMY-2 and bla
CTX-M-15 resistance genes as well as for the IncFIB and IncI1 replicons, at 42°C for 18 h with subsequent washings, as recommended by the manufacturer. Purified PCR products were used as hybridization controls and E. coli C600 and E. coli J53 (plasmid-free strains) as negative controls (Supplementary Fig. S1; Supplementary Data are available online at
Results
E. coli isolates identified
CTXR E. coli isolates were detected in 11 of 71 cloacal samples analyzed (15.5%), which corresponded to 10 pet turtles and one environmental habitat turtle. Three CTXR isolates were initially selected from each positive sample, however, after bacterial identification, susceptibility testing, and assays of PCRs for the identification of β-lactamase genes, some isolates looked similar; for this reason, only one isolate per sample was maintained and further studied, doing a collection of 11 CTXR E. coli isolates.
Detection of ESBL-positive E. coli isolates and clonality
Four of 11 CTXR E. coli isolates showed a positive ESBL screening test, which were recovered from two females and two males of two different turtle species (Table 1). The gene encoding CTX-M-2 enzyme was identified in two of them and the gene of CTX-M-15 in the other two isolates (Table 1).
Conjugative strains as well as beta-lactamases and plasmids transferred by conjugation are indicated in bold letters. Beta-lactamases and associated plasmids detected by Southern blot hybridization are indicated in underlined letters.
Sex: F, female; M, male; Origin previous to herpetarium shelter: P, pet; E, environment.
No ST complex assigned.
bla CTX-M-15 gene and the type of plasmid where it was localized by Southern blot hybridization.
bla CMY-2 gene and the type of plasmid where it was localized by Southern blot hybridization.
ND, non determined.
ESBL, extended-spectrum β-lactamases; MLST, multilocus sequence typing; PFGE, pulsed-field gel electrophoresis; ST, sequence type.
The CTXR E. coli strains were classified in nine different PT by the cluster analysis of Dice similarity index (value >85%) (Fig. 1). Two groups of CMY-2-producing isolates (PT8 and PT9) showed a 100% Dice coefficient of clonal identity, as well as the same ST2329 and ST155, respectively (Fig. 1).
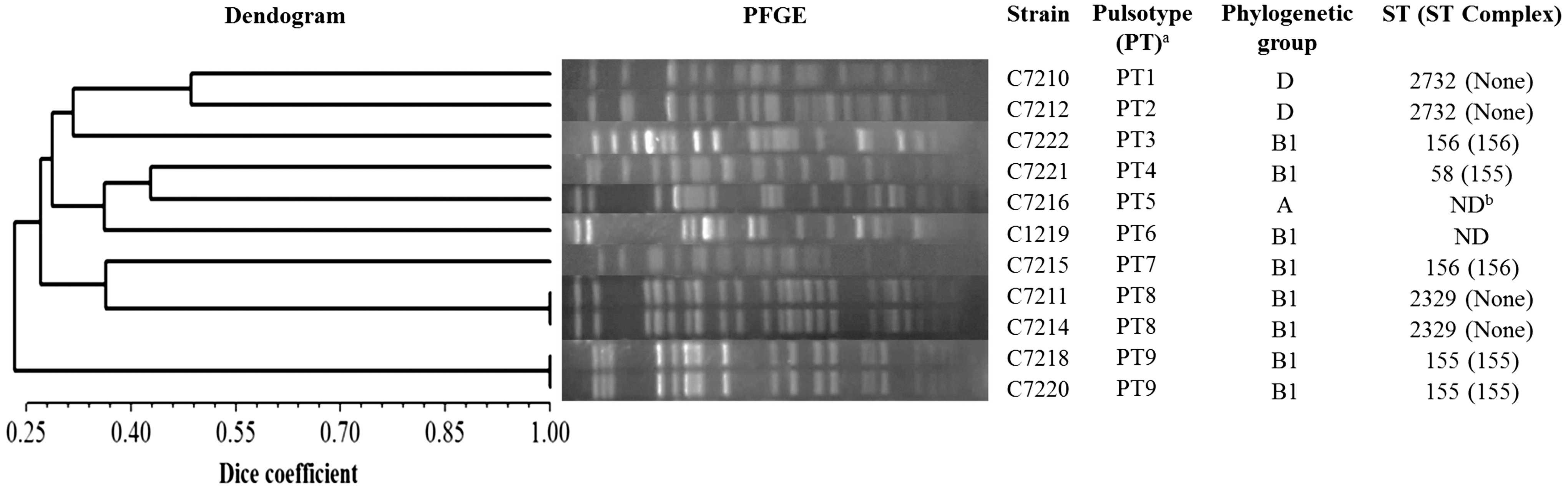
UPGMA dendogram, PFGE of XbaI-digested genomic DNA patterns, and molecular typing from the 11 ESBL and CMY-2-producing Escherichia coli strains of turtle origin. aClassification based on the cluster analysis of Dice similarity index (value >85%). bNon determined. ESBL, extended-spectrum β-lactamases; PFGE, pulsed-field gel electrophoresis; UPGMA, Unweighted Pair Group Method using arithmetic averages.
Detection of acquired pAmpC β-lactamase-producing E. coli isolates
Eight of the 11 CTXR E. coli isolates showed resistance to amoxicillin–clavulanic acid and/or cefoxitin and 7 of them exhibited a negative ESBL screening test. The CMY-2 β-lactamase was identified by PCR and sequencing in all these eight isolates (one of them also carried the bla CTX-M-2 gene showing a positive ESBL screening test) (Table 1). The bla CMY-2 gene was flanked by the ISEcp1 and tnpA in upstream and by the blc gene in downstream regions, as determined by PCR and sequencing of flanking regions.
The CMY-2-positive E. coli isolates showed a high genetic diversity with six different PFGE PTs; these isolates were ascribed to the phylogroups B1 (n = 6), A (n = 1), and D (n = 1), and to the sequence types ST155, ST156, ST2329, and ST2732. Different mutations in the ampC-regulatory region were identified in six CMY-2-positive isolates (−18, −1, +58 in all these isolates, with an additional change in +54 in two of them) (Table 2).
NA, nalidixic acid; CI, ciprofloxacin; G, gentamicin; T, tobramycin; S, streptomycin; C, chloramphenicol; TM, trimethoprim; SXT, trimethoprim/sulfamethoxazole; TE, tetracycline. The breakpoints for the antimicrobial agents were compared with CLSI (2013).
Resistance determinants transferred by conjugation to the E. coli recipient strains C600 or J53 are indicated in bold letters.
These structures correspond to those represented in Figure 2.
ND, non determined.
ni, non identified array with performed PCRs.
Antimicrobial resistance genes and integrons
Table 2 shows the resistance genes detected in the 11 CTXR E. coli isolates. Five strains carried plasmid-mediated quinolone resistance genes, detected by PCR and sequencing [qnrA, qnrB19, aac(6′)Ib-cr, and oqxB]. The CMY-2-positive E. coli strain C7222 harbored the three genes qnrB19, aac(6′)Ib-cr, and oqxB. The tetracycline resistance tet(A) and/or tet(B) genes were detected in 10 of 11 ESBL/pAmpC-E. coli isolates (except in strain C7212). At least three amino acid changes were identified in GyrA (two changes at positions 83 and 87) and ParC (from one to two changes at positions 80 and 84) of the five ciprofloxacin-resistant E. coli strains (Table 2).
Class 1 and 2 integrons were characterized by the primer walking strategy and sequencing. The intI1 gene was detected in 9 of 11 ESBL/pAmpC isolates, and five different gene cassette arrays were identified (Fig. 2, Table 2). Four of these arrays corresponded to nonclassical integrons lacking the conserved 3′-CS region (qacEΔ1 and sul1 genes), most of them linked to sul3 gene. The fifth gene cassette array corresponded to a classical class 1 integron with the 3′-CS region, and the array dfrA1-aadA1. Four E. coli strains showed a positive PCR for intI2 gene, and the array dfrA1-sat2-aadA1 was identified in two of them (corresponding to CTX-M-2-positive isolates) (Fig. 2, Table 2).

Structures of gene cassette arrays detected in class 1
Gene transfer assays and plasmid analysis
At least three different replicons were identified by PCR in the 11 ESBL/pAmpC-E. coli isolates of the study, being IncFrep, IncFIB, IncK, and IncI1 the most prevalent replicon plasmids detected (Table 1). The E. coli isolates carried from two to four plasmids ranging in sizes from 45 to 220 kb, determined by S1-PFGE strategy (Fig. 3).
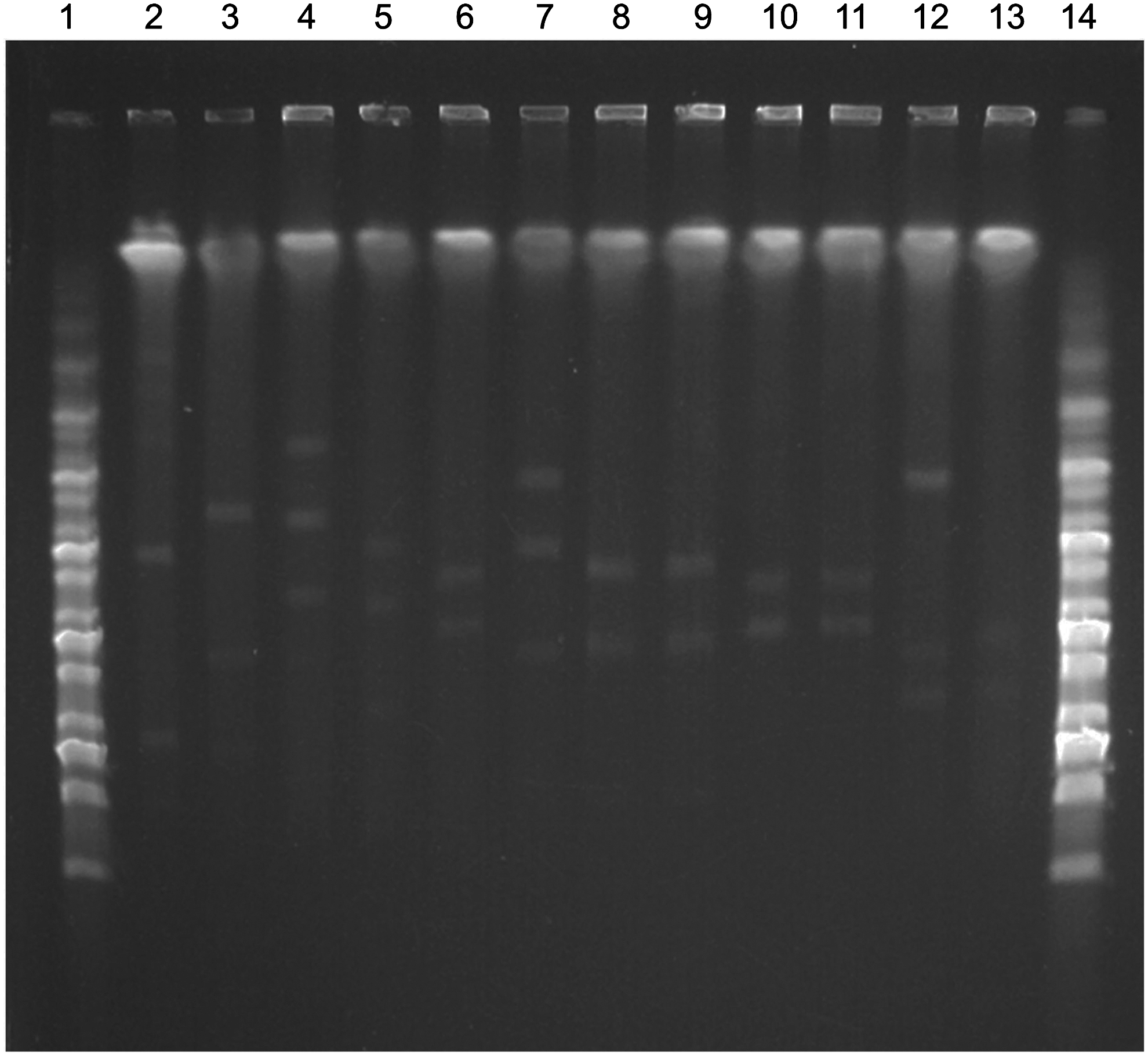
PFGE of Nuclease S1-digested DNA to determine the number and size of plasmids from the 11 ESBL or CMY-2-producing E. coli strains of turtle origin. 1 and 14, MidRange I PFG Marker (New England Biolabs®); 2, plasmid-containing reference strain NCTC 50192 (showing approximated plasmids of 154, 66, and 48 kb); 3, C7210, 4, C7212; 5, C7221; 6, C7215; 7, C7222; 8, C7211; 9, C7214; 10, C7218; 11, C7220; 12, C7219; 13, C7216.
Conjugation experiments were performed in 9 of the 11 ESBL/pAmpC producing E. coli strains, which presented unrelated PFGE patterns. Transconjugants were obtained from four of these nine isolates with two recipient strains: (1) strain C7221 (recipient E. coli C600); (2) strains C7214, C7219, and C7222 (recipient RIFR J53 E. coli).
The bla CTX-M-15 gene was localized by hybridization in an IncFIB plasmid of 120 kb in the conjugative strain C7221 (Table 1), as well as in its transconjugant (Table 1 and see Supplementary Figs. S1 and S2. This ESBL gene was also detected in a plasmid of 135 kb in the nonconjugative strain C7215 (Table 1 and Supplementary Fig. S2). In contrast, the bla CMY-2 gene was detected in IncI1 plasmids of 85 kb (conjugative strain 7222) or 95 kb (strain C7216), as well as in an IncFIB plasmid of 90 kb in the conjugative strains C7214 and C7219 (Table 1 and Supplementary Figs. S1 and S2). Moreover, the bla CMY-2 gene of the strains C7218 and C7220 was localized in a 125 kb plasmid, although it could not be determined its specific replicon type; both strains presented the same sequence type (ST155) and PFGE pattern (PT9) and were recovered from two turtles of the same species (T. scripta elegans). During the conjugative experiments, cotransference of qnrA and bla CMY-2 genes was evidenced in C7219 strain (Tables 1 and 2).
Discussion
Beta-lactamases of the ESBL or pAmpC types are frequently detected in E. coli isolates worldwide, being the CTX-M-15- and the CMY-2-type enzymes the most prevalent in human isolates (Cantón et al. 2012); these enzymes inactivate third generation cephalosporins, and they can mediate resistance to carbapenems, when combining with other resistance mechanisms (Bortolaia et al. 2014).
The high frequency of detection of ESBL- and especially of pAmpC-producing E. coli strains among the studied turtles (15.5%) is of special relevance. Although our E. coli isolates presented a high diversity of PFGE patterns (nine different PTs) and STs (five different ones), a partial clonal dissemination could not be discarded, due to the fact that some of the animals carried E. coli isolates with identical or related PFGE patterns (C7211/C7214, C7218/C7220, and C7210/C7212) (Fig. 1), which could contribute, in part, to the high prevalence detected. Nevertheless, the dissemination of plasmids among different E. coli clones of the intestinal tract of these turtles is a plausible possibility (Chen et al. 2010).
The origin of these ESBL/pAmpC-producing E. coli isolates might be related to the food used for these animals (chicken liver, vegetables, mealworms, or commercial turtle biscuits), since ESBL- and AmpC-producing E. coli have been extensively reported in food-producing animals, especially in chickens (Ferreira et al. 2014). Nevertheless, we cannot exclude the potential acquisition of these multidrug-resistant bacteria during the preherpetarium period (either in-house as pets or in wildlife in the environment), mostly if we consider that most of the studied turtles came from the household environment before they were sheltered in the herpetarium.
The ESBL types detected in turtles, CTX-M-2 and CTX-M-15, correspond to variants frequently reported among clinical E. coli isolates in Latin America (García-Fulgueiras et al. 2011, Sennati et al. 2012, Redondo et al. 2013) and Mexico (Rocha-Gracia et al. 2010, Silva-Sánchez et al. 2013), respectively.
The CTX-M-15 variant was detected in isolates of lineages ST58 and ST156, as previously reported in E. coli isolates from humans and animals in The Netherlands and Chile (Dierikx et al. 2012, Hernandez et al. 2013); however, the lineage ST2732 associated to CTX-M-2 β-lactamase as in this study, has not been previously reported. Moreover, the pandemic clone B2-ST131 was not found in our collection of strains.
It is of relevance the high frequency of CMY-2-producing isolates among the studied healthy animals (8 of 71, 11.3%). CMY-2 is the most common pAmpC β-lactamase worldwide disseminated in clinical isolates (Jacoby 2009). Moreover, the CMY-2 variant has been detected in Salmonella isolates from humans and E. coli isolates from healthy dogs in Mexico (Wiesner et al. 2013, Zaidi et al. 2013, Rocha-Gracia et al. 2015) and in South America (Cejas et al. 2014). It is noteworthy that Salmonella is associated with colonization in reptiles (Chen et al. 2010, Whitten et al. 2015), as turtles (Díaz et al. 2006, Liu et al. 2013); for this reason, a possible transference of bla CMY-2 gene between Salmonella and E. coli isolates could not be discarded. It has been previously demonstrated the transference of common plasmids carrying bla CMY-2 gene between animal-associated Salmonella and E. coli isolates, and the presence of these plasmids also in human isolates. All these facts suggest that plasmid-mediated CMY-2 β-lactamase has undergone transference between different bacterial species and hosts (Winokur et al. 2001), causing a serious concern of public health due to the growing popularity of animal reservoirs as pets and the close contact with humans. The presence of CMY-2 E. coli producers isolated from turtles that were used as pets represents an evolutionary advantage for the spread of β-lactam resistance determinants as a consequence of the high use of β-lactam antibiotics in both human and veterinary medicine (Bortolaia et al. 2014).
It is interesting that the phylogroup B1, which has been associated with commensal and environmental isolates, was the most frequently identified among E. coli isolate producers of resistance enzymes as CTX-M-15 or CMY-2 β-lactamases. On the other hand, the CTX-M-2 E. coli producers belonged to the phylogroup D, which has been associated to extraintestinal pathogenic strains, with more virulence factors than commensal E. coli isolates of phylogroups A or B1 (Carlos et al. 2010).
The bla CMY-2 gene has been associated with IncI1, IncA/C, IncFII, IncI2, IncL/M, and IncB/O plasmids in E. coli isolates from humans, companion and food animals (Bortolaia et al. 2014, Guo et al. 2014). Our strains contained different replicons as is the case of IncFIB, IncI1, IncFrep, and IncK as the most prevalent ones (Table 1), and the bla CMY-2 gene was detected in IncI1 or IncFIB plasmids of 85–95 kb in four of them (in two of these strains the ESBL encoding plasmid was transferred by conjugation). The size of the plasmids carrying the bla CMY-2 gene (125 kb) was detected in two of the strains, but not the plasmid replicon type. The bla CMY-2 gene was also cotransferred with qnrA gene in C7219 strain, but neither gene was localized in the 195 kb IncFIB plasmid detected by hybridization in the donor strain. The plasmid characterization suggests a high diversity of conjugative and nonconjugative plasmids harboring the bla CMY-2 gene among turtle E. coli isolates in this study. The possible exchange of plasmids harboring bla CMY-2 between humans and animals is mostly focused to studies examining bacterial isolates from human, food-producing animals, and food (Winokur et al. 2001, Guo et al. 2014), and the spread of this β-lactam resistance determinant from pets has received less attention to date. In this study, the genetic diversity observed in plasmids encoding CMY-2 represents an evolutionary advantage that could facilitate spread of this β-lactam resistance determinant, with implications in both human and veterinary medicine.
In addition, a variety of genetic elements have been implicated in the mobilization of pAmpC encoding genes, like the insertion sequence ISEcp1, associated with bla CMY-2, which is able to mobilize and supply an efficient promoter for the high-level expression of neighboring genes, like the chromosomal bla gene onto plasmids (Jacoby 2009). The ISEcp1 element was found in the genetic environment of blaCMY-2 gene in pAmpC-producing E. coli isolates of our study.
The global spread of CTX-M-15 ESBL type is mainly associated with incompatibility group F plasmids, although IncN, IncI1, and IncL/M plasmids are also involved (Cantón et al. 2012); we detected an IncFIB plasmid of 120 kb carrying the bla CTX-M-15 gene in a conjugative strain and in its transconjugant. The bla CTX-M-15 gene was also in a nonconjugative plasmid of 135 kb that did not hybridize with IncFIB or IncI1 probes; we cannot discard that this plasmid corresponded to IncK type, but specific hybridization was not performed.
The studied animals contained fluoroquinolone-resistant E. coli isolates (7%), with PMQR genes associated. The presence of oqxB gene is very unusual in E. coli, although it has been previously reported from isolates of swine manure source in Denmark (Hansen et al. 2004), farm animals, farm workers and the environment in China (Zhao et al. 2010), as well as in a clinical E. coli isolate in Spain (Ruiz et al. 2012), among other sources. This efflux pump encoding gene (commonly found as a chromosomal gene in Klebsiella pneumoniae), which is associated with a reduction in fluoroquinolone susceptibility, was found coassociated with other PMQR genes in a CMY-2-producing E. coli strain of lineage ST156 and different plasmid replicons, fact that could explain the extended genetic quinolone-resistant background.
Furthermore, the presence of sulfonamide, chloramphenicol, and trimethoprim resistance genes was detected in class 1 or class 2 integrons (most in arrays associated to non-classical integrons) as previously reported (Sáenz et al. 2010). This mechanism could be implicated in the dissemination of resistance determinants among these ESBL/pAmpC-producing E. coli isolated from turtles.
Conclusions
To our knowledge, this is the first report of ESBL, plasmid-encoded AmpC β-lactamases, PMQR genes and mobile determinants of antimicrobial resistance in E. coli of turtle origin. The presence of both CTX-M 15 (global epidemic variant) and CTX-M-2 (more associated to the American continent) β-lactamases as well as the plasmid-encoded CMY-2 β-lactamase is of interest because healthy turtles may be a source of E. coli producing these enzymes and could participate in the dissemination of multidrug-resistant bacteria to other environments and hosts. Future studies are necessary to determine the mutual dissemination of multidrug-resistant bacteria between turtles used as pets and humans, upon clinical, veterinary, and ecological environments; this fact is of relevance considering that nowadays turtles have become very popular pets that have direct contact with humans, especially with children, highlighting the role of turtles as a possible close source of multidrug-resistant bacteria.
Footnotes
Acknowledgments
The authors thank Escuela de Biología of Benemérita Universidad Autónoma de Puebla for access of animals included in this study. This study was approved and supported by the Vicerrectoría de Investigación y Estudios de Posgrado, of the Benemérita Universidad Autónoma de Puebla across the Programa Institucional de Fomento a la Investigación y a la Consolidación de Cuerpos Académicos, Sub-programa de Aseguramiento de Investigadores Consolidados, convocatorias 2012, 2013 y 2014. Also, it was supported for CONACYT-México, across the summons of Ciencia Básica 2012-01, with number of reference 178942. G.C.C. has a fellowship of CONACYT with number 233611 and C.A.A. has a predoctoral FPI fellowship of the Ministerio de Economía y Competitividad of Spain (BES-2013-063105). The work performed in University of La Rioja was financed by project SAF2012-35474 from the Ministerio de Economía y Competitividad of Spain and the Fondo Europeo de Desarrollo Regional (FEDER). The authors thank Eduardo Miguel Brambila Colombres for the use of his laboratory for hybridization experiments. Part of these results has been presented at the 24th European Congress of Clinical Microbiology and Infectious Diseases, Barcelona, 10th–13th May, 2014.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.