
Abstract
The co-evolution of Orientia tsutsugamushi and its vector/host Leptotrombidium mites is important for this bacterium to survive and exist in its environment. The data in this study demonstrated that O. tsutsugamushi has adapted to take advantage of the parasitic nature of the host's larval stage and thus increase its chance of transmission to a vertebrate host and potentially to other vector mites by increasing its density at the time of transmission. Our data demonstrated that during the larval stage the density of O. tsutsugamushi was at its highest level compared to other life stages (Kruskal–Wallis, p < 0.0001). We further revealed that the different O. tsutsugamushi 56-kDa type-specific antigen (TSA) genotypes within the mite were maintained and preserved during transovarial transmission from the Leptotrombidium imphalum, lines Li-3 and Li-5. No sequence difference of 56-kDa TSA gene (variable domain I–III, 765 bp) was observed between the UT302-like genotype found in mothers and their offspring (100% identity). However, one or two nonsynonymous mutations in the 56-kDa TSA gene were observed in the Karp-like genotypes found in the F1 offspring with a percent difference ranging from 0.13 to 0.26 for nucleotide sequences and from 0.39 to 0.78 for amino acid sequences. Additionally, the composition of co-existing O. tsutsugamushi genotypes was maintained in L. imphalum lines through transsovarial and transstadial transmission processes; however, the proportion of these genotypes in each stage varied (larva, nymph, adult). These results show some of the key characteristics of O. tsutsugamushi maintenance within and transmission among its vector/host L. imphalum.
Introduction
O
Earlier studies of electron microscopy of O. tsutsugamushi in infected mites showed a unique electron-dense outer layer of its cell wall structure, which differs markedly from that of members of the Rickettsia (Silverman and Wisseman 1978). This unique characteristic of the cell envelope was identical in all forms of O. tsutsugamushi found within infected mite tissues. Three forms of O. tsutsugamushi were observed, a typical form with dimensions similar to the Karp strain infecting L-929 cells, an elongate form with a bundle of microtubules at the pole, and an electron-dense or pleomorphic form (Wright et al. 1984). Typical forms were generally found in the salivary glands of the larval stage, whereas electron-dense (pleomorphic) and elongate forms were mainly found in the digestive epithelium of the gastric cecae and ovarian tissue in adult stage, respectively. Generally, all stages of an infected mite were infected, but the location of bacteria within the mite shifted from one organ to others during the various development stages, resulting in an observation of bacteria accumulating in the salivary gland cells and mid gut in the larval stage and in ovarian tissues in the adult mite (Roberts et al. 1975, Wright et al. 1984, Kadosaka and Kimura 2003). The localization of O. tsutsugamushi in particular organs in larval and adult stages is consistent with the transmission of bacteria from vector to host or with the transovarial transmission to its offspring. In addition, the bacteria distributions in 7-day and 60-day-old adult mites were similar, but the older females were generally more heavily infected.
Mice inoculation experiments showed that O. tsutsugamushi could be isolated from all life stages of infected L. fletcheri besides the parasitic larval stage of the mite (Roberts et al. 1975). Several studies have supported the presence of bacteria during the stages of development from larva to nymph to adult, but other studies have indicated that the bacterium was not found within eggs of infected female adults right after oviposition; rather they were detected in various parts of the embryo before the hatching, suggesting a bacterial multiplication process occurred during embryogenesis (Urakami et al. 1988). This observation was supported by the results published by Kollars et al. (2001), showing that the infection rate in eggs produced by infected L. imphalum colonies was very low, with a percent infection ranging from 8.1% to 45.6%.
Vertical transmission of O. tsutsugamushi in Leptotrombidium mites has been considered to be an efficient strategy for bacteria to be maintained in all successive generations during its life cycle (Frances et al. 2001). Contrarily, horizontal transmission does not appear to be an efficient method because the orientiae were rarely found to be acquired by uninfected larvae while feeding on infected mice (Frances et al. 2000). On the other hand, efficient vertical transmission has been demonstrated in several studies as well as in laboratory-reared Leptotrombidum mite colonies (Phasomkusolsil et al. 2009). An infected L. chiangraiensis has been found to convey O. tsutsugamushi 100% of the time by transovarial and filial transmission in two successive generations studied. However, an infected L. imphalum colony has been found to be a little less efficient, with 100% and 93.3% transovarial transmission in parental and F1 generation, respectively, and 100% and 62.3% filial transmission in F1 and F2 generations, respectively. Moreover, the symbiosis of O. tsutsugamushi in L. deliense mites did not affect the number of eggs produced from female adults and the time of development from engorged larva to adult (Frances et al. 2001).
Our previous study showed the co-existence of two O. tsutsugamushi genotypes in single laboratory-reared mite lines (Takhampunya et al. 2014). Twelve lines from three species of Leptotrombidium mite—L. chiangraiensis (Lc), L. imphalum (Li), and L. deliense (Ld)—were studied. An O. tsutsugamushi Karp-like genotype was found either as a single genotype as in the L. chiangraiensis lines Lc-1, Lc-2, Lc3, and Lc-5 or as a co-infection with an O. tsutsugamushi UT302-like genotype found in the L. imphalum lines Li-1, Li-2, Li-3, Li-4, Li-5, Li-6, and Li-7, or with the O. tsutsugamushi Gilliam-like genotype in the L. deliense line Ld-1. The co-existing O. tsutsugamushi genotypes were found in all larvae in L. imphalum and L. deliense colonies according to our previous data using three larvae randomly picked from each line. In the present study, the relative density of O. tsutsugamushi in all development stages was monitored as well as the variation and composition of genotypes during transovarial and transstadial transmission process.
Materials and Methods
Laboratory-reared O. tsutsugamushi–infected mites
Seven O. tsutsugamushi–infected L. imphalum mite lines were derived from seven infected females. These mite lines are maintained in an animal biosafety level-3 (ABSL-3) facility at the Armed Forces Research Institute of Medical Sciences (AFRIMS) (Table 1) (Takhampunya et al. 2014). Larval mites fed upon the ears of International Cancer Research (ICR) mice from a Charles River Technology (BioLASCO, Taiwan) colony maintained by the Department of Veterinary Medicine, AFRIMS, whereas nymphs and adults were fed a diet of springtail eggs (protocol # 09-11, Maintenance of the Leptotrombidium larval mite colonies: Chigger feeding on ICR mice [Mus musculus]). In this study, three L. imphalum lines (Li-2, Li-3, Li-5) (Takhampunya et al. 2014) were used in all experiments. Samples of L. imphalum Li-2 line stages larva, nymph, and adult (Fig. 1) were collected at different time points, depending on the availability of samples during February to July, 2014. Figure 2 shows the relationship of specimens collected from the L. imphalum Li-2 line in relation to the infected parental L. imphalum Li-2 chigger (P1) collected from a Rattus rattus host in Chiangrai Province in 1993 (Takhampunya et al. 2014).

Images of the L. imphalum Li-2 line at different development stages (larva, nymph, and adult). Pictures were taken with a Nikon Eclipse microscope at 400× (Nikon, Japan).
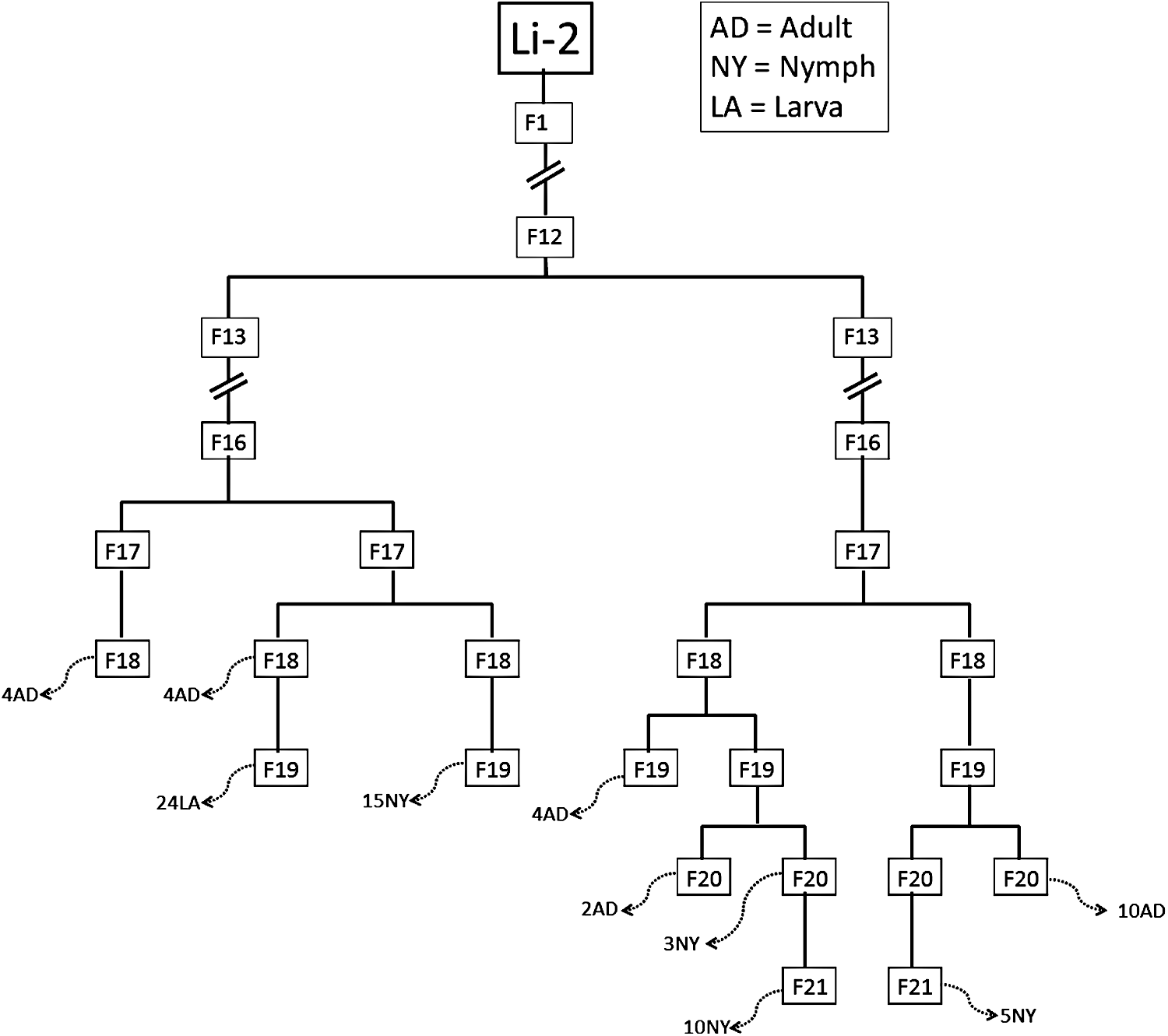
Specimens used in this experiment were collected from multiple filial passages of the O. tsutsugamushi–infected L. imphalum Li-2 line during February to July of 2014. Twenty-four adults (AD), 33 nymphs (NY), and 24 larvae (LA) were collected from several filial passages with the sampling numbers indicated before the capital letters (broken arrows). Li-2 refers to the original infected chigger (parent) collected from R. rattus in 1993 and her successive offspring through several generations from F1 to F21. Three specimens of each stage assessed selected for determining the composition of O. tsutsugamushi genotypes as shown in Table 2.
Extraction of genomic DNA from O. tsutsugamushi–infected mites
O. tsutsugamushi–infected mites from three different lines of L. imphalum were subjected individually to genomic DNA extraction using a modified tissue protocol from a QIAamp® DNA Mini Kit (Qiagen, Hilden, Germany) and a previously published protocol (Takhampunya et al. 2014).
O. tsutsugamushi densities in individual Leptotrombidum mites
The quantitative real-time PCR (qPCR) assay, Otsu47 qPCR assay (Jiang et al. 2004), was used as previously described, except that the probe was replaced with SYBR Green to detect and quantitate the presence of the O. tsutsugamushi 47-kDa HtrA gene in individual mites. The Leptotrombidium small-subunit rRNA (ssrRNA) gene (12S rRNA) qPCR assay, using SYBR Green, was conducted to determine the densities of Leptotrombidium DNA in individual mite samples and used as reference gene for normalization of qPCR data. In-house primers, ssrRNA_F (5′-caaatggcgtgccagcagtctcgg-3′) and ssrRNA_R (5′-aaacttctactttgttacgacttacc-3′), were designed from ssrRNA region of Leptotrombidium scutellare (GenBank acc. no. AB300497). In every mite sample, two qPCR assays, the Otsu47 qPCR assay and the Leptotrombidium 12S rRNA assay, were performed at the same time. The reaction mixture (30 μL) contained 15 μL 2× SYBR Green qPCR SuperMix-UDG (Invitrogen), 0.2 μM each of primers, and 1 μL (47 kDa), or 0.5 μL (ssrRNA) of DNA template. The qPCR assay was performed by incubating the sample at 50°C for 2 min, 95°C for 2 min, followed by 35 cycles of 95°C for 15 s and 60°C for 1 min. Each 96-well plate contained triplicate reaction mixtures for each DNA sample (individual mite), the appropriate set of standards, no-template controls, and negative controls (uninfected mites).
Melt curve analysis was performed following the completion of the real-time PCR cycles to analyze the identity of the product. All reactions were performed with the ABI 7500 Fast Real-Time PCR System (Life Technologies, Carlsbad, CA). Ten-fold dilutions of calibration standards (O. tsutsugamushi 47-kDa HtrA gene and Leptotrombidium 12S rRNA gene) were detectable down to 100 copies of all DNA targets. Standard curve slopes, correlation coefficients, and amplification efficiencies were calculated using ABI 7500 System SDS software version 1.4.1 (Life Technologies), and the results are shown in Supplemental Table 1 (Supplementary Data are available at
Amplification and sequencing of the 56-kDa type-specific antigen gene
A fragment of the 56-kDa type-specific antigen (TSA) gene was amplified by nested PCR (nPCR) using previously described primers (Horinouchi et al. 1996) and in-house primers designed from published sequences (Takhampunya et al. 2014). The detailed protocol, amplification conditions, and primer sequences were published previously (Takhampunya et al. 2014).
Cloning and sequencing of the O. tsutsugamushi 56-kDa TSA gene
The PCR product of a fragment of the 56-kDa TSA gene (765 bp) was purified by a Qiagen QIAuick PCR Purification Kit (Qiagen), cloned into a pCR2.1®-TOPO® Vector (Invitrogen, Foster City, CA), and grown in Escherichia coli (DH5α™-T1R). Depending on each experiment, a certain number of E. coli colonies carrying a plasmid successfully harboring the 56-kDa insert were selected randomly for sequencing. Plasmids were purified using a QIAprep Spin Miniprep Kit (Qiagen) and sent for DNA sequencing at AITBiotech (Singapore).
Characterization of the O. tsutsugamushi genotype by PCR-restriction fragment length polymorphism assay
The PCR product of the 56-kDa TSA gene (765 bp) was cut with the restriction endonuclease enzyme HhaI (Biolabs, UK) for 2 h at 37°C to distinguish the O. tsutsugamushi genotypes from the infected mite line Li-2. Genotype controls for the Li-2 infected line were generated from cloning of the 56-kDa TSA PCR product into a TOPO Vector (Invitrogen). The genotypes were confirmed by DNA sequencing.
Sequence and phylogenetic analysis
Sequences were edited and assembled using Sequencer program version 4.1.4 (Applied Biosystems) and compared to sequences in the BLASTN database (National Center for Biotechnology Information). Multiple sequence alignment was performed using the ClustalW program (Thompson et al. 1997). The neighbor-joining (NJ) tree with bootstrapping (2000 replicates) was constructed using the p-distance model of nucleotide substitution (56-kDa gene variable domains I–III) in the MEGA 6 program (Tamura et al. 2013). Synonymous and nonsynonymous nucleotide substitution rates were computed using the joint maximum likelihood approach (Muse and Gaut 1994) and Tamura–Nei model of nucleotide substitution (Tamura and Nei 1993) in the MEGA 6 program.
Statistical analyses
The difference of O. tsutsugamushi densities between stages was tested by the Kruskal–Wallis test, and the critical range (p < 0.001) was determined. All statistical calculations were performed with GraphPad Prism® version 5.04 software (La Jolla, CA).
Animal care and use
The experiments reported herein were conducted in compliance with the Animal Welfare Act and in accordance with the principles set forth in the “Guide for the Care and Use of Laboratory Animals,” Institute of Laboratory Animals Resources, National Research Council, National Academy Press, 1996.
Results
Maintenance of O. tsutsugamushi–infected Leptotrombidium mites in an ABSL-3 laboratory
Samples were collected from L. imphalum Li-2, Li-3, and Li-5 lines for each stage of its life cycle—larval, nymph (deutonymph), and adult—for a period of 6 months. All specimens were disinfected with −70°C ethanol in a glass vial before transfer to the laboratory for analyses. The representative pictures of larval, nymph, and adult stages of Li-2 line are presented in Figure 1. The size of the mites increased substantially as they transformed from six-legged larva to eight-legged nymph, and to adult.
The variation of O. tsutsugamushi density measured during the life stages of L. imphalum mites
To examine and monitor the O. tsutsugamushi density during the different development stages of Leptotrombidium mites, 24 larvae, 33 nymphs, and 24 female adults were collected from a laboratory-reared O. tsutsugamushi–infected L. imphalum Li-2 line. Figure 2 shows the relationship of specimens collected from L. imphalum Li-2 line in relation to the infected parental L. imphalum Li-2 chigger (P1) collected from a R. rattus host in Chiangrai Province in 1993 (Takhampunya et al. 2014). qPCR assays of the O. tsutsugamushi 47-kDa gene (Otsu47) and Leptotrombidium 12S rRNA (mite housekeeping gene) were used to compare the density of O. tsutsugamushi in each stage. The density was calculated and the data from individual mites of each group/stage were plotted and compared against each other. The density of O. tsutsugamushi in the larval stage is significantly higher than adult and nymph stages (Kruskal–Wallis, p < 0.0001) by 2.9 and 4.2 orders of magnitude, respectively (Fig. 3). The adult stage had a mean O. tsutsugamushi density higher than the nymph stage but without statistical significance (Kruskal–Wallis, p > 0.05). Huge variations of O. tsutsugamushi densities were observed in larval stage specimens, with density ranging from 0.015 to 0.096 (mean, standard deviation [SD] = 0.046 ± 0.025), whereas the range of O. tsutsugamushi density in nymph and adult stages was relatively small, with density ranging from 0.001 to 0.046 (mean, SD = 0.011 ± 0.011) in nymphs and from 0.004 to 0.039 (mean, SD = 0.016 ± 0.011) in adults.
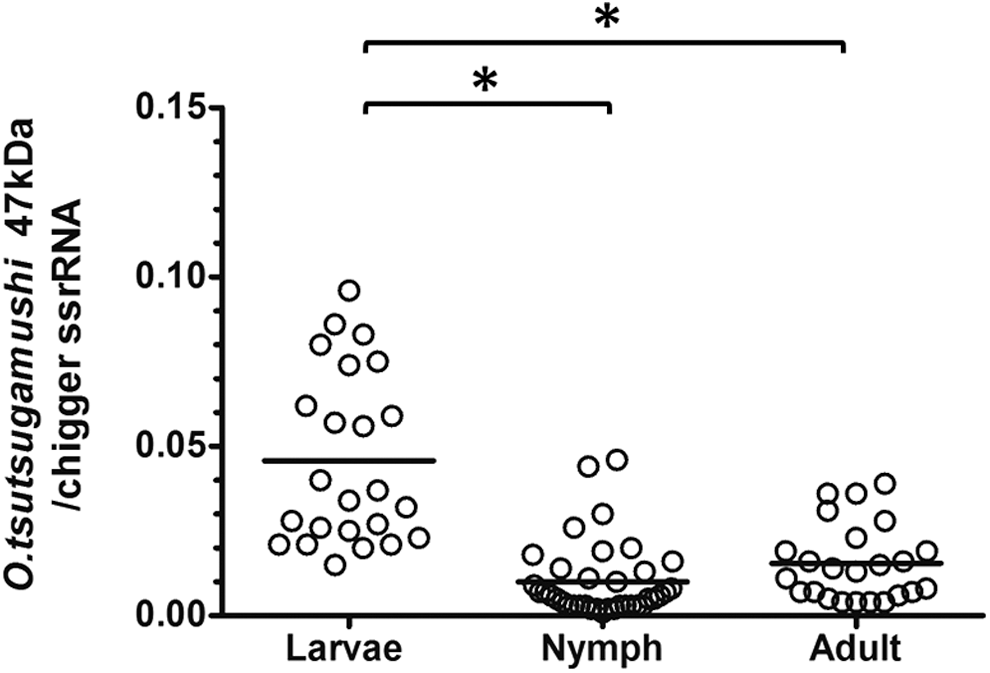
Variation of O. tsutsugamushi densities in different developmental stages of L. imphalum Li-2 line. O. tsutsugamushi densities in individual laboratory-reared L. imphalum Li-2 mites were measured. A dot plot graph displays the densities O. tsutsugamushi as determined by qPCR assays of O. tsutsugamushi 47-kDa gene per the Leptotrombidium 12S rRNA gene. The solid line shows mean of bacterial densities; larva (mean, standard deviation [SD] = 0.046 ± 0.025, n = 24), nymph (mean, SD = 0.011 ± 0.011, n = 33), female adult (mean, SD = 0.016 ± 0.011, n = 24). Asterisks (*) indicate the statistical significance at p < 0.0001.
The composition of O. tsutsugamushi genotypes through the different development stages of Leptotrombidium mite (transstadial transmission of co-existing O. tsutsugamushi genotypes)
Our previous published data showed that the L. imphalum Li-2 line consists of two O. tsutsugamushi genotypes: One is Karp-like and the other is UT302-like (Takhampunya et al. 2014). For this study, the composition of these two genotypes during the development stages was studied. A PCR-restriction fragment length polymorphism (RFLP) assay was applied to determine the O. tsutsugamushi genotype using the 56-kDa TSA gene as the target. The patterns of RFLP generated from specimens were compared with genotype controls. As shown in Figure 4, the controls generated different patterns, as shown in lane 1 (UT302-like) and lane 2 (Karp-like). Almost all specimens from larvae, nymphs, and adults had a mixture of these two RFLP patterns, suggesting that the two genotypes co-exist in single L. imphalum Li-2 mites during various life stages. Table 1 summarizes the results of genotyping of O. tsutsugamushi by PCR-RFLP assay in 24 larvae, 33 nymphs, and 24 female adults. All larvae (100%) and adults (100%) consist of mixed genotypes (Karp-like and UT302-like); however, not all nymphs assessed (97.0%) had mixed genotypes. One nymph (3.0%, N2 lane in Fig. 4) had only the Karp-like genotype.
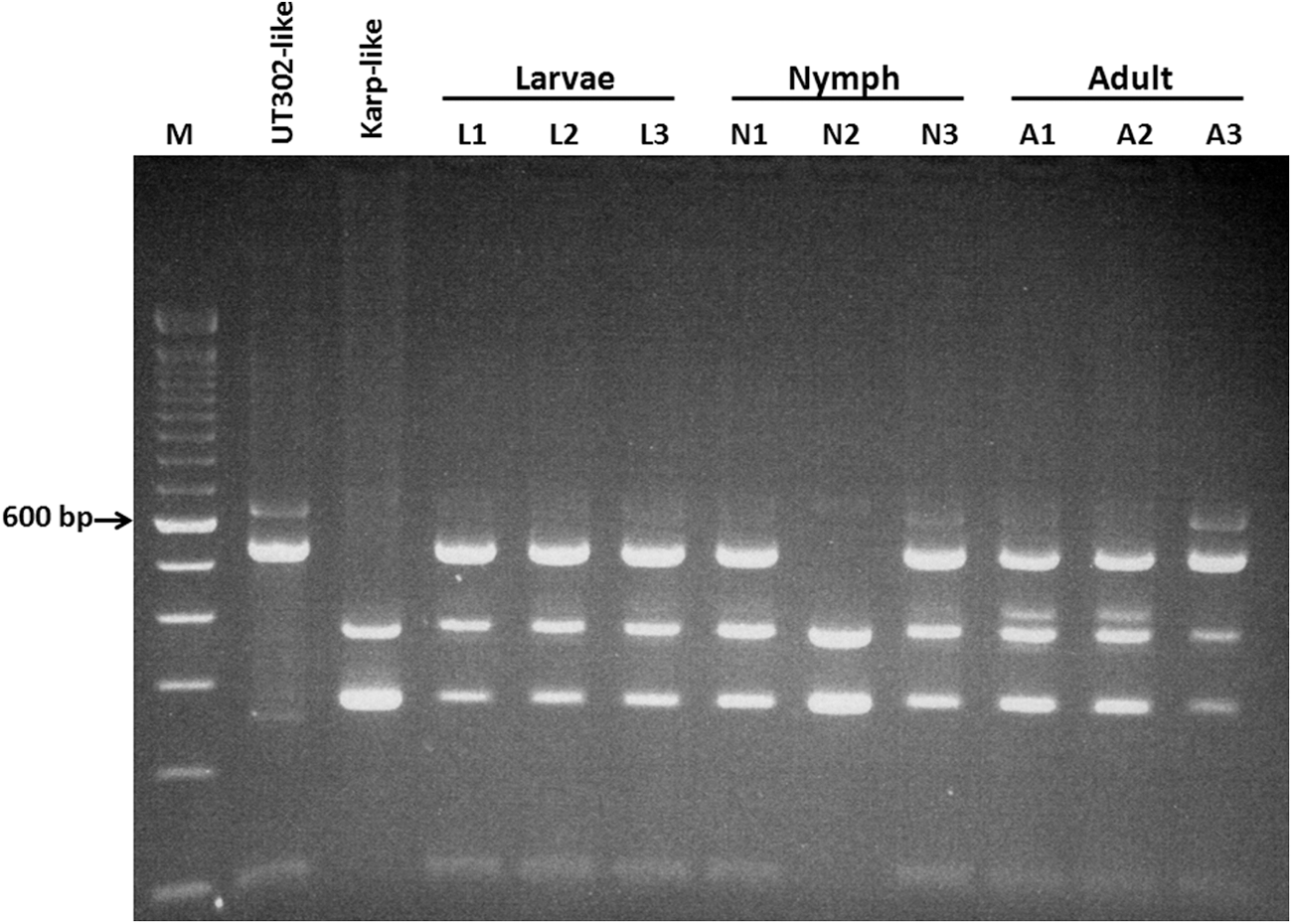
O. tsutsugamushi genotypes in L. imphalum Li-2 line using a PCR-restriction fragment length polymorphism (RFLP) assay. Representative RFLP patterns found in larvae, nymphs, and adults are shown. Positive controls for UT302-like and Karp-like are shown in lanes 1 and 2, respectively. Larvae (lanes L1–L3), nymphs (lanes N1–N3), and adults (lanes A1–A3) are indicated. Lane M contains molecular markers.
The proportion of O. tsutsugamushi Karp-like to UT302-like genotypes in individual mite was investigated in three specimens from each stage (larva, nymph, and adult). Only specimens infected with two genotypes were selected for this experiment. Each PCR product of the 56-kDa gene amplified from these mites was cloned into a TOPO vector, as described previously (Takhampunya et al. 2014). Twenty to 22 transformed colonies containing the 56-kDa–inserted vector were randomly picked from nine specimens. The genotype in each plasmid was determined by PCR-RFLP, as mentioned above. The results of genotyping are shown in Fig. 4, larval and adult stages have a proportion of UT302-like genotypes to Karp-like genotype of around 6:4, whereas nymph showed an opposite result of a 4:6 ratio of UT302-like to Karp-like genotypes (Table 2).
Three samples of each stage (larva, nymph, and adult) were studied. Specimens were derived from multiple filial passages of the L. imphalum Li-2 line (see also Fig. 2). In each sample, 20–22 cloned colonies were picked and genotyped by PCR-RFLP assay as shown in Fig. 4.
SD, standard deviation.
Transovarial transmission of co-existing O. tsutsugamushi genotypes in Leptotrombidium mites
The composition of O. tsutsugamushi genotypes in infected Leptotrombidium mites during transovarial transmission was investigated and is presented here. Female adult mites of two infected L. imphalum lines (Li-3 and Li-5) and two of their F1 offspring (larval stage) were examined. DNA sequencing of the O. tsutsugamushi hypervariable region (domains I–III) of the 56-kDa TSA gene was performed. Ten bacteria colonies with the correct inserted fragment of 56-kDa gene PCR product (∼765 bp) were randomly picked and subjected to DNA sequencing as described in Materials and Methods section. As shown in Table 3, O. tsutsugamushi in an L. imphalum Li-3 mother (Li-3 Mo) consists of UT302-like and Karp-like genotypes at a ratio of 9:1 (UT302-like: Karp-like). Two larvae from its F1 generation (Li-3 F1#1 and F1#2) had the ratios of 8:2 and 6:4 (UT302-like: Karp-like), respectively. In the L. imphalum Li-5 mother (Li-5 Mo), the ratio of UT302-like to Karp-like was 1:1, and its F1 generations had 1:1 (Li-5 F1#1) and 7:3 (Li-5 F1#2). From this data, the composition and proportion of two genotypes were maintained comparatively during transovarial transmission.
For each mite specimen, 56-kDa TSA PCR amplicons were cloned into TOPO vectors, and 10 randomly picked transformed bacterial colonies were selected for DNA sequencing and analyses.
To demonstrate if the bacteria have changed through the transovarial transmission process, the diversity of O. tsutsugamushi genotypes was investigated in mother mites and their larvae (F1 generation). The DNA sequences of 56-kDa TSA gene fragments from the above experiment were analyzed. Multiple sequence alignments and phylogenetic analyses were conducted to examine their sequence diversity and relationships with reference genotypes. As shown in NJ tree in Figure 5, the 60 sequences from mother mites (Li-3 Mo, Li-5 Mo) and their F1 generations (Li-3 F1#1, F1#2 and Li-5 F1#1, F1#2) fall into two major groups: Group 1 with two patterns is within the UT302 genotype (94.1–100% identity), and group 2 with two patterns is within the diverse Karp-like genotype (98.3–100% identity) (Fig. 6). The majority of group 1 UT302-like sequences (97.5%, 39/40 sequences) detected in mother mites of Li-3 Mo clone 1-9, Li-5 Mo clone 1-4, and their F1 larvae; Li-3 F1#1 clone 1-8, Li-3 F1#2 clone 1-6, Li-5 F1#1 clone 1-5, and Li-5 F1#2 clone 1-7 are identical with 100% identity found among them (pattern 1). However, a sequence from Li-5 Mo clone 5 (mother mite) is unique (pattern 2), having only 94.1% identity to the other 39 UT302-like sequences found in Li-3 and Li-5 mites. Further analyses revealed the significant differences of this pattern 2 sequence from the rest of UT302-like sequences. There were 36 mutations in nucleotide sequences, five synonymous and 31 nonsynonymous, resulting in 17 amino acid changes (6.6%) between pattern 2 and pattern 1 sequences. Additionally, three amino acid insertions (1.2%) were observed, two amino acids (six nucleotides) at one site and one amino acid at another site.

Neighbor-joining (NJ) tree constructed from partial 56-kDa TSA genes (variable domain I–III, 765 bp) of O. tsutsugamushi. The analysis involved 60-nucleotide sequences of O. tsutsugamushi–infected L. imphalum Li-3 and Li-5 lines; mother mite (Mo) and two F1 offspring (F1#1 and F1#2), along with 32 reference sequences retrieved from the GenBank database. Ten partial 56-kDa TSA gene sequences of randomly picked bacterial colonies from six individual mites were included in this analysis, as indicated by clone 1 to clone 10 following each sample name. The NJ tree was constructed using the p-distance method of nucleotide substitution model. The bootstrap p values >50% are shown (2000 replicates). The tree is rooted at its midpoint. Scale bar represents substitution per site. The expanded data set for group 2 and Karp-like genotypes (shading area) is shown in Fig. 6.

Subtree of O. tsutsugamushi Karp-like genotypes and group 2 from Fig. 5 (shaded area) consisting of sequences detected from L. imphalum Li-3 and Li-5 lines. Percent nucleotide and amino acid differences are shown in Table S2.
Among the 20 Karp-like sequences ascertained from the L. imphalum Li-3 and Li-5 mite lines, two patterns were recognized. The first pattern (pattern 3) consisted of seven Karp-like sequences from the Li-3 line samples (Li-3 Mo clone 10, Li-3 F1#1 clone 9-10, and Li-3 F1#2 clone 7-10), which grouped together with 100% identity, except for one sequence of Li-3 F1#1clone 9 with a 99.9% identity (Fig. 6). This Li-3 F1#1 clone 9 had one nucleotide mutation (nonsynonymous) resulting in one amino acid change (0.39%). Thirteen Karp-like sequences identified among the Li-5 line 9 sequences grouped with pattern 3 consisted of samples from Li-5 Mo clone 8-10, Li-5 F1#1 clone 7-10, and Li-5 F1#2 clone 9-10, with a percent identity ranging from 99.6–100%. These include two sequences of Li-5 F1 #1 (clone 9-10) and F1#2 (clone 9-10) that were identical to their mother (Li-5 Mo clone 9-10), whereas the Li-5 F1#1 clone 7 sequence is more similar to a second Li-5 Mo clone 8 with 99.7% identity (two-nucleotide mutations, nonsynonymous) and a Li-5 F1#1 clone 8 sequence that had only one one nucleotide mutation (nonsynonymous) when compared with the two different Li-5 Mo clones (clone 8, clone 9-10) (Fig. 6). A second pattern (pattern 4) of Karp-like sequences was recognized from two Li-5 Mo clones (clone 6 and clone 7) and a single clone from Li-5 F1#1 clone 6 and F1#2 clone 8, with a percent identity ranging from 99.7% to 100%. The two Karp-like sequences from Li-5 F1#1 clone 6 and Li-5 F1#2 clone 8 were identical to Li-5 Mo clone 6 (Fig. 6). Collectively these data show that the three patterns of genotypes found in F1 larvae are acquired from the mother mites, and the sequence diversity within these three genotype patterns is very limited.
Discussion
O. tsutsugamushi, an obligate intracellular parasite, survival depends upon its host, the Leptotrombidium mites. Thus, its dynamic bacterial density and localization within the host complies with the orientiae's needs to survive and exist throughout the host's biological developmental changes. Correspondingly, we measured higher bacteria densities in the parasitic and active stage of the Leptotrombidium mites (larval stage), resulting in an enhanced chance for the orientiae to be transmitted to other hosts. Similar observations were noted by others, such as the accumulation of orientiae in active organs of each life stage, i.e., in salivary glands of the larval stage and ovarian tissues of the female adults (Roberts et al. 1975, Wright et al. 1984). Moreover, it was found that the bacteria density in the nymph stage was less than that found in other developmental stages (i.e., larva and adult).
It is possible to speculate that during development from larva to nymph (active to quiescent period) the O. tsutsugamushi residing in the host cells remain inactive or with minimal activity, resulting in lower bacteria density. These observations are supported by the previous study of Roberts et al. (1975) showing that in an infected L. fletcheri colony the mouse isolation rate of O. tsutsugamushi from unfed, engorged larvae and nymphophanes was higher than the isolation rates from nymphs and adults. The bacteria were most commonly found in the salivary glands and mid gut of larvae and in the ovaries of the female adults, whereas the bacteria in the nymphs were distributed in all organs but at a lower rate. Thus, our data and those of others suggest that the activities of the orientiae are directly linked to the host mites' developmental changes, potentially to the bacterium's benefit.
In our previous study, we demonstrated co-existing O. tsutsugamushi genotypes in single larva (Takhampunya et al. 2014). The data from this study are supportive of that earlier investigation and now show that the co-infection of two genotypes in a single mite is maintained in the mite lines throughout the biological development of the mite, and these genotypes are passed on to the mite's offspring. In addition, this study demonstrated how co-existing genotypes were maintained during transovarial and transstadial transmission in terms of proportion and diversity of the O. tsutsugamushi.
Transovarial and transstadial transmissions are very efficient and consistent processes for O. tsutsugamushi to be maintained in and co-evolve with its mite host. All larvae, adults, and 97% of nymphs (32/33) assessed maintained the mixed infection of Karp-like and UT302-like genotypes. However, the proportion of these two genotypes in each stage (larva, nymph, adult) varied, although the proportion was generally maintained during the transovarial transmission to the offspring (F1). The explanation behind this variable proportion event is unknown, although it has been described previously (Shirai et al. 1982). In the Shirai et al. report, the authors used a direct fluorescent antibody (FA) assay to investigate the number and identity of O. tsustusgamushi serotypes maintained within L. fletcheri and L. arenicola lines of their mite colony. Their results indicated that naturally infected mite colonies retained multiple serotypes of O. tsutsugamushi through successive generations; however, their detection by the FA assay varied from familial line to line and from generation to generation, even though the chiggers maintained the same serotypes from generation to generation.
Genetic investigation of the mixed O. tsutsugamushi agents determined that there was limited diversity of the 56-kDa TSA (including three of four variable domains) gene observed during the transovarial transmission, with 100% identity of UT302-like genotype found in both the mother mites and their successive F1 larvae and over 99.7% identity for Karp-like genotypes in mother mites and their offspring (maximum of two nucleotides difference). There was one exception that is worth noting. The Li-5 Mo clone 5, UT302-like sequence, was unique (not found in the mother mite's progeny or other mites assessed), and it was extremely dissimilar from the other UT302-like genotype (94.1% identity). However, when additional progeny and mites were investigated by targeted gene sequencing using the GS Junior System (Roche 454 Life Sciences, Branford, CT), this particular genotype was detected reproducibly (data not shown).
Although two genotypes were found to co-exist and appeared to have co-evolved within a single host, there was no evidence of new or unrelated sequences found in the F1 larvae, implying that recombination events within the 56-kDa TSA gene fragment assessed between the two genotypes had not occurred during single filial passage. However, this conclusion about recombination events is only applicable to the 765-bp fragment containing the I–III variable domains of the 56-kDa TSA gene. There could be sequence variation occurring in other regions (e.g., IV variable domain) of this gene as well as other genes. The ability of mites to maintain a specific genotype over many successive generations has been shown previously (Kelly et al. 1994). These authors showed that a unique O. tsutsugamushi genotype, as determined by RFLP analysis of the 22-kDa antigen gene, was found to be maintained for 60 generations in a familial line of L. fletcheri over a 26-year period.
It has been shown that the multiplication of O. tsutsugamushi occurs during the development of an embryo in mite eggs (Urakami et al. 1988) and in older female adults (Wright et al. 1984), allowing the bacteria to increase its abundance in the host so that they can survive into the next generation of the host. Moreover, the accumulation of bacteria was shown to shift from one location/organ to another (active organ) during the various developmental stages of the mite life cycle (Roberts et al. 1975), suggesting that the bacteria have evolved and adapted to the host biology and behaviors so that they can survive and co-exist with the host. In conclusion, we have confirmed that not only do individual O. tsutsugamushi genotypes co-evolve with their Leptotrombidum mite hosts, but that more than one O. tsutsugamushi genotype can co-evolve with Leptodrombidium mites and that they are able to keep their genotype identity preserved.
Footnotes
Acknowledgments
We are grateful for Mr. Taweesak Monkanna and Mr. Opas Thachinin managing the mite colony and in providing mite specimens for this study. This work was supported by the DoD MIDRP program work unit # 6000.RAD1.WJ.A0310.
Author Disclosure Statement
No competing financial interests exist. The views expressed in this article are those of the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, nor the U.S. government. The author, A.L.R., as an employee of the U.S. government, conducted this work as part of his official duties. Title 17 U.S.C. § 105 provides that “copyright protection under this title is not available for any work of the United States government.” Title 17 U.S.C § 101 defines a U.S. government work as a work prepared by an employee of the U.S. government as part of the person's official duties.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.