
Abstract
Background:
Apart from infection with human filariae, zoonotic filariasis also occurs worldwide, and the numbers of cases have been increasing steadily. Diagnosis of intact filariae in tissues or organs depends on histological identification. The morphology of parasites in tissue-embedded sections is poor and shows high levels of homoplasy. Thus, the use of morphological characteristics in taxonomic studies is difficult and may not allow a specific diagnosis.
Methods:
Here we report the use of real-time PCR with high-resolution melting analysis (HRM) to detect and identify Brugia malayi, Brugia pahangi, Wuchereria bancrofti, and Dirofilaria immitis in paraffin-embedded sections. Assay specificity was determined using other tissue-dwelling parasites, Angiostrongylus cantonensis, Gnathostoma spinigerum, and Cysticercus cellulosae. We also developed a quick paraffin removal protocol.
Results:
Both human and animal filariae in formalin-fixed paraffin-embedded sections (FFPES) were diagnosed and identified rapidly, whereas other parasites were negative. There was no difference in the melting temperature of products amplified from filarial DNA obtained from unstained FFPES and Hematoxylin & Eosin–stained sections. Therefore, the DNA extraction protocols developed in this study could be used for real-time PCR with HRM.
Conclusions:
We report the successful application of a HRM-PCR assay to differentiate four filarial parasites in FFPES, thus providing the pathologist with an effective alternative diagnostic procedure. Furthermore, the quick paraffin removal protocol developed could shorten the duration and number of steps required for paraffin removal using a standard protocol.
Introduction
F
An unusual presentation of filariasis with a large number of microfilariae in subcutaneous swelling was previously reported (Garbyal et al. 2008). Furthermore, humans have been reported to be infected with zoonotic filariae including Brugia pahangi and Dirofilaria species (Dissanaike 1979), and recently the number of cases has increased steadily worldwide (Orihel and Eberhard 1998). Many of these zoonotic infections are now more widely recognized by clinicians and pathologists (Pampiglione et al. 1995). The reasons for the increasing number of cases may be due to increased exposure to zooanthropophilic mosquito bites. Some mosquitoes carry human-specific filariae, whereas others carry larvae from animal filariae. Human dirofilariasis caused by infection with Dirofilaria worms has been frequently reported. Since the first report of dirofilariasis in Hungary in 1879 (Szenasi et al. 2008), the prevalence of human dirofilariasis has increased (Kramer et al. 2007).
Zoonotic filaria infections are typically cryptic (Simmons et al. 1984, Nozais et al. 1994). A case of filariasis as the cause of gynecomastia clinically mimicked carcinoma because the host tissue response to the parasites resulted in a hard breast lump. Thus, pathologists should screen cytological smears carefully, especially those from patients from endemic areas, to exclude coexistent parasitic pathology for a timely diagnosis and appropriate management (Kohli and Jain 2012).
A definitive diagnosis of intact parasites in human tissue is made through pathological examination of the tissue specimens. However, in most cases, finding parasitic structures is difficult because the parasite morphology in tissue-embedded sections is often poor with high levels of homoplasy. Furthermore, species identification is often difficult if the intact parasite lacks key characteristic morphology (Orihel and Eberhard 1998, Morales-Hojas, 2009). Additionally, not all intact filariae are well described and characterized.
Molecular-based diagnostic tools have been used in laboratories for reliable differential identification of filarial parasites. Many studies have shown good results for detection and identification of lymphatic filariae by molecular biology-based methods, such as PCR-linked restriction fragment length polymorphism (PCR-RFLP) (Nuchprayoon et al. 2005, Nuchprayoon et al. 2006), multiplex PCR (Latrofa et al. 2012), and real-time PCR with high-resolution melting curve analysis (HRM) (Wongkamchai et al. 2014). The advantages of HRM are that amplification and detection steps can be conducted in the same reaction tube or well, and no post-PCR processing is required (Dobrowolski et al. 2009, Nettuwakul et al. 2010). The master mix used in HRM contains a DNA-binding dye that stains target sequences and results in sharp melting signals that are analyzed using Gene Scanning Software in the HRM instrument. (Dobrowolski et al. 2009). The assay can be run rapidly with large sample sets, such as 96 or 384 samples, in one PCR run, with a duration of approximately 5 min for melting and analysis. The cost effectiveness of HRM is comparable to or cheaper than conventional PCR and sequencing (Zhou et al. 2005, Dobrowolski et al. 2009).
A recent application of HRM for diagnosis and differentiation of filarial infection was demonstrated (Pangasa et al. 2009, Adaszek et al. 2010, Ngui et al. 2012, Ceccarelli et al. 2014, Wongkamchai et al. 2014). We targeted the mitochondrial partial 12S rRNA gene when developing real-time PCR with HRM analysis for differentiation of filarial parasites because it is highly conserved and contains genus- and species specific sequence variations useful for identifying filaria species, i.e, Brugia malayi, B. pahangi, and Dirofilaria immitis (Wongkamchai et al. 2013).
Thus, molecular technology may be an alternative method for the diagnosis of lymphatic filarial infection in formalin-fixed paraffin-embedded sections (FFPES). Formalin fixation followed by paraffin embedding of tissue specimens is a widely used preservation method because it helps to maintain morphological features of the tissue specimen. Nevertheless, the use of HRM real-time PCR with FFPE samples to identify parasitic infection has not been reported.
The objectives of the present study were to: (1) Determine whether HRM-PCR is a useful method for identification and differentiation of filarial infection in FFPE specimens; (2) develop a rapid paraffin removal method and compare it with the standard paraffin removal technique; and (3) test the interference of Hematoxylin & Eosin (H&E) staining in DNA extraction by comparing it with unstained FFPE specimens.
Materials and Methods
Ethical approval
This study was conducted according to the Helsinki Declaration and was approved by the Ethical Committee of the Faculty of Medicine, Siriraj Hospital, Mahidol University, based on the Ethics of Human Experimentation of the National Research Council of Thailand. The Certificate of Approval number is Si285/2013.
Study samples
Study samples consisted of adult stages of B. malayi, B. pahangi, and D. immitis were provided from TRS Labs, Athens, Georgia, and an archived histopathological tissue specimen contained structures of the female stage of a lymphatic filarial worm located in enlarged lymph node tissue was provided from Department of Pathology, Faculty of Medicine Siriraj Hospital, Mahidol University. The adult stage of Angiostrongylus cantonensis, third stage larva of Gnathostoma spinigerum, and Cysticercus cellulosae purchased from the Faculty of Tropical Medicine, Mahidol University were used for a cross-reactivity study. Positive controls for the HRM-PCR assay consisted of DNA from B. malayi, B. pahangi, D. immitis, and Wuchereria bancrofti.
Preparation of formalin-fixed paraffin-embedded sections
Adult stages of B. malayi, B. pahangi, D. immitis, A. cantonensis, the third-stage larva of G. spinigerum, and C. cellulosae were fixed in 10% formalin to preserve tissues and maintain the cell and subcellular component structures. For dehydration, samples were transferred through progressively increasing alcohol concentrations (80–95%) and absolute alcohol (three times/h). Samples were then transferred through xylene (three times/h) for clearing (Prophet et al. 1992).
For embedding, samples were transferred through paraffin (three times/h) and soaked in paraffin under vacuum for 1 h. A processing cassette containing an impregnated tissue was removed from the paraffin holding area, and a mold that best corresponded to the size of the tissue sample was selected and partially filled with paraffin. The sample was removed, placed at the bottom of the mold, and transferred from a warm plate to a cold plate for further sectioning. Before sectioning, excess paraffin was trimmed, leaving a margin at the top and bottom of the block. For rough cutting, the block was thick sectioned by repeatedly advancing the block manually and slicing. Next, the knife and block were chilled using an ice cube and sections cut at a slow and even speed. Paraffin sections were lowered into a water bath until flat, then separated and placed onto clean premarked glass slides that were drained vertically for several min before being placed onto a warming table at 37–40°C overnight. Then FFPES were kept at room temperature until used for further DNA extraction or staining.
H&E staining
For H&E staining, sections were deparaffinized in xylene for 30 min followed by incubation in 100% ethanol for 10 s. Sections were rehydrated by incubation in a graded ethanol series (80%, 60%, and 40%) for 10 s each and finally distilled water for 10 s. Slides were stained with Mayer's Hematoxylin solution (Dako, Santa Clara, CA) for 15 min and washed under running tap water for 15 min. Slides were then incubated in distilled water, placed in 80% ethyl alcohol for 1–2 min, counterstained in Eosin-Phloxine solution (Dako, Santa Clara, CA) for 2 min, dehydrated, and cleared through two changes each of 95% ethyl alcohol, absolute ethyl alcohol, and xylene for 2 min each. Stained slides were mounted using resinous medium.
DNA extraction from FFPES using different paraffin removal protocols
FFPES were divided into three groups. Group 1 included unstained FFPES prepared from adult stages of B. malayi, B. pahangi, D. immitis, A. cantonensis, third-stage larva of G. spinigerum, and C. cellulosae. This group used a standard paraffin removal protocol prior to DNA extraction. Briefly, unstained FFPES were incubated in xylene for 30 min followed by a graded ethanol series from 100% ethanol (dehydration) (80%, 60%, and 40%) for 10 s each and rehydrated by using double-distilled water for 10 s. Then, 200 μL of tissue-lysis buffer was added onto the slide. Sections were scraped off the glass slide and transferred to a 1.5-mL Eppendorf tube, followed by DNA extraction with High Pure PCR Template Preparation Kit for tissues according to the manufacturer's protocol (Roche Diagnostics,).
Group 2 consisted of unstained FFPES prepared from adult stages of B. malayi, B. pahangi, D. immitis, A. cantonensis, third-stage larva of G. spinigerum, and C. cellulosae. The quick paraffin removal protocol prior to DNA extraction was used in this group. Glass slides of unstained FFPES were placed on a hot plate for 1 min, then placed vertically to remove melted paraffin, followed by quick addition of 200 μL of tissue-lysis buffer onto the slide and scraping the section into a 1.5-mL Eppendorf tube. DNA extraction was by High Pure PCR Template Preparation Kit protocol (Roche Diagnostics).
Group 3 contained H&E-stained FFPES prepared from adult stages of B. malayi, B. pahangi, D. immitis, A. cantonensis, third-stage larva of G. spinigerum, and C. cellulosae as well as an archived histopathological tissue specimen. First, 200 μL of tissue-lysis buffer was dropped onto the slide. The stained section was scraped off the slides and transferred to a 1.5-mL Eppendorf tube. DNA extraction was by High Pure PCR Template Preparation Kit protocol (Roche Diagnostics). The extracted DNA concentration from all groups was measured by a Nanodrop spectrophotometer (Thermo Fisher Scientific Waltham, MA) according to the manufacturer's instructions. DNA was used for the HRM-PCR assay.
Primers for B. malayi, B. pahangi, D. immitis, and W. bancrofti amplification
Previously designed primers targeting the mitochondrial partial 12S rRNA gene, which is highly conserved and contains genus- and species-specific sequence variations (Wongkamchai et al. 2013), were used for identifying the four filaria species (B. malayi, B. pahangi, D. immitis, and W. bancrofti). Figure 1 shows the primers based on alignment of the mitochondrial partial 12S rRNA genes of B. malayi (GenBank acc. no. AJ544843; positions 177–287), B. pahangi (AM779851; positions 154–264), D. immitis (FN391554; positions 161–275), and W. bancrofti (NC_016186.1; positions 7619–7729).

Alignment of partial mitochondrial 12S rRNA gene sequences from B. malayi (GenBank acc. no. AJ544843; positions 177–287), B. pahangi (AM779851; positions 154–264), D. immitis (FN391554; positions 161–275), and W. bancrofti (NC_016186.1; positions 7619–7729). Dots indicate identity, dashes indicate deletion from the above consensus sequence, and grey areas indicate primers.
Real-time PCR with HRM analysis for detection and identification of filarial species
Real-time PCR assay was performed for species detection and identification in the paraffin-embedded unstained/H&E stained sections of adult stages of B. malayi, B. pahangi, and D. immitis and third-stage larva of G. spinigerum, adult stage of A. cantonensis, and C. cellulosae and an archival histological tissue section. The real-time-PCR mixture contained 5 μL of commercial ready-to-use reaction mix for PCR containing LightCycler 480 ResoLight Dye (Roche Diagnostics), 10 μmol/L of each primer, and 2 μL of DNA extract with nuclease-free water added to a final volume of 10 μL. Each set of experiments included negative and positive controls. All PCR reactions were carried out in a 96-well clear PCR plate for LightCycler 480 II (Roche Diagnostics) with nontemplate controls. The reaction was performed in a thermal cycler as follows: 95°C for 5 min (for polymerase activation), followed by 40 cycles of 95°C for 10 s (denaturation); 58°C for 10 s (annealing); and 72°C for 10 s (extension). Melting curves were generated, normalized, temperature-shifted, and converted to difference plots by LightCycler® 480 gene scanning software (Roche Diagnostics).
DNA sequencing
DNA extracted from the archival histological tissue section was amplified using conventional PCR and submitted for DNA sequencing.
Results
HRM-PCR assay
Photomicrographs of Hematoxylin & Eosin (H&E) stained FFPES prepared from filarial parasites and other tissue-dwelling parasites (Fig. 2) as well as from an adult female filarial parasite from tissue section of a cervical lymph node of a patient (Fig. 3).
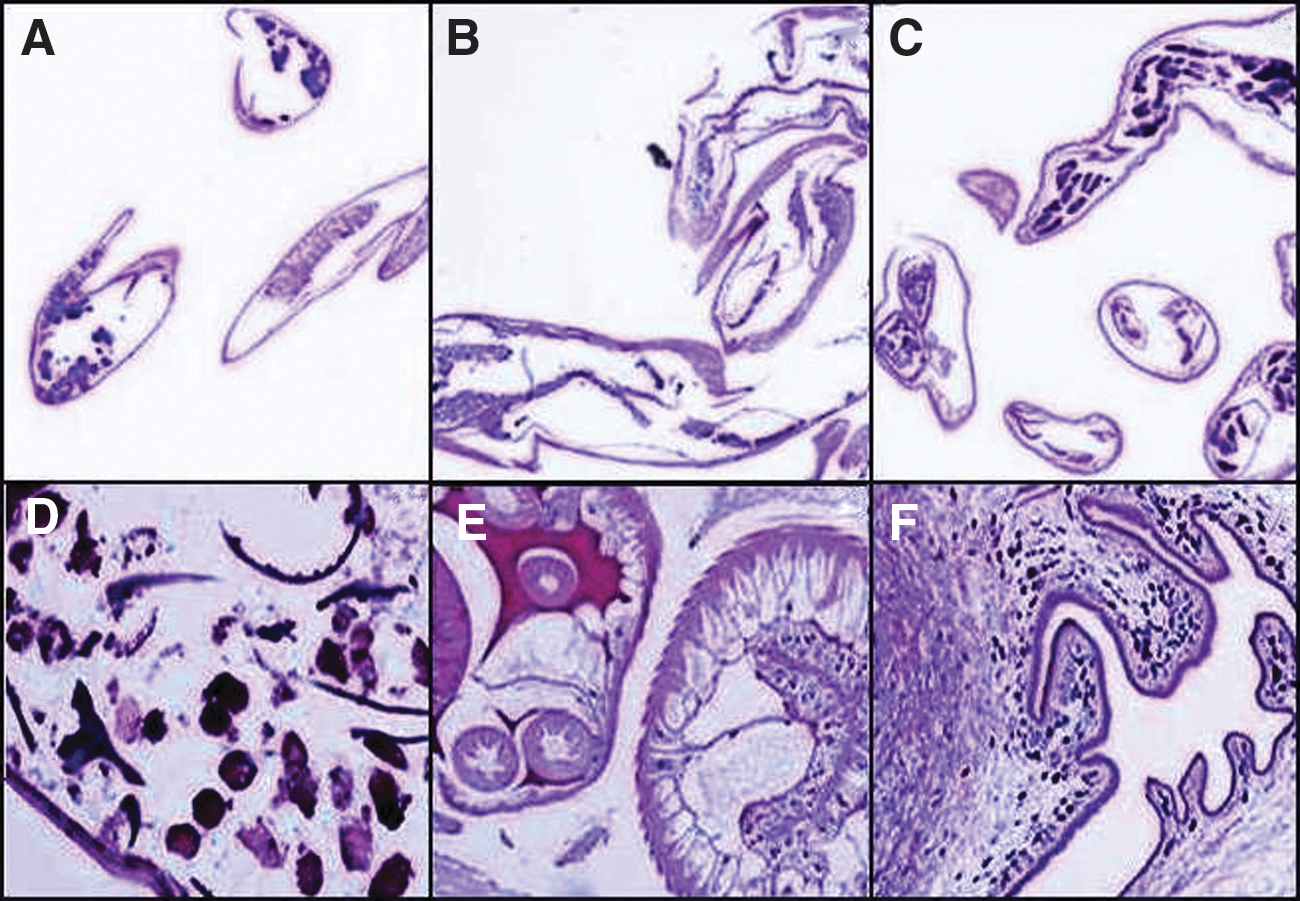
Photomicrographs showing Hematoxylin & Eosin (H&E)-stained transverse sections of three filarial parasites (
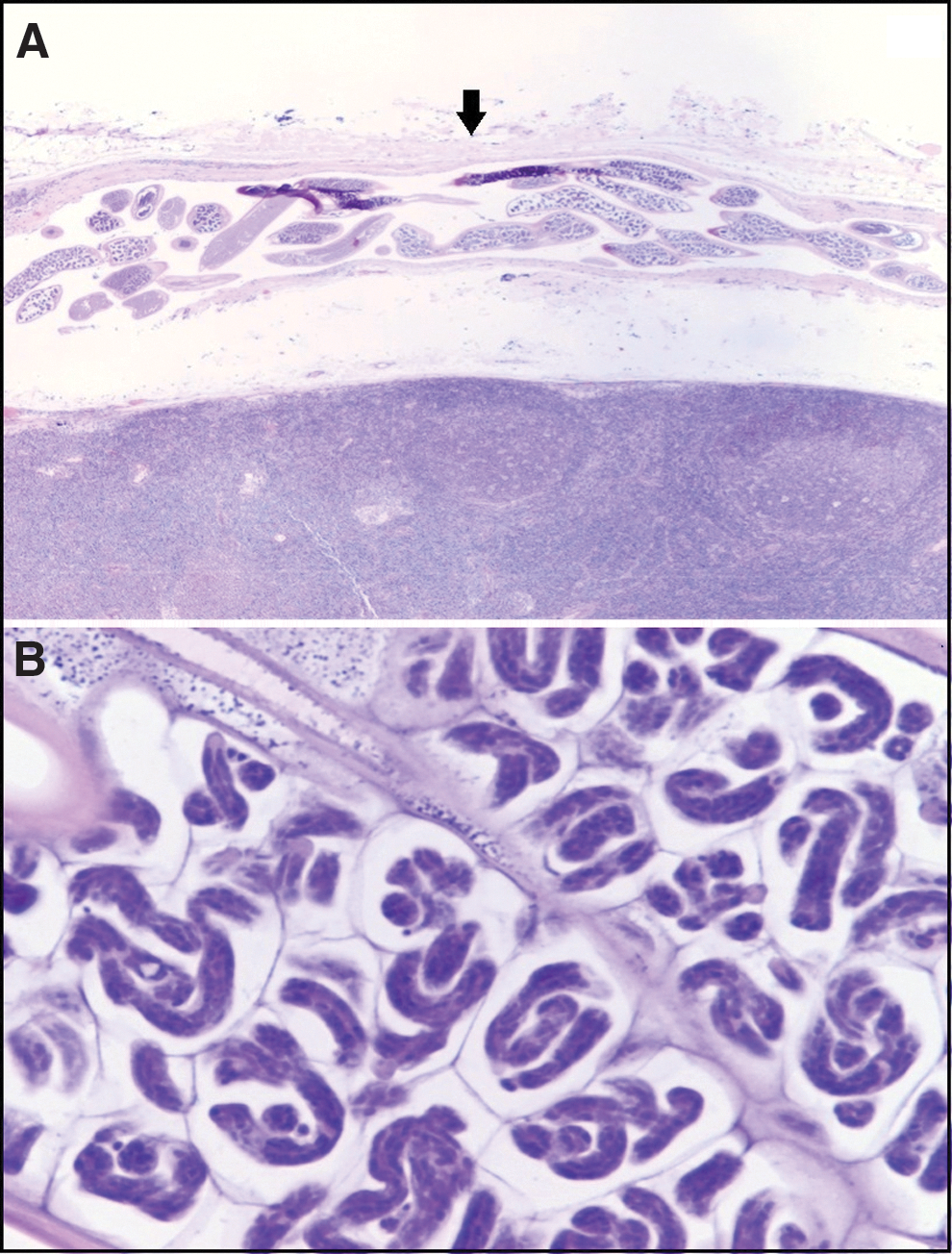
Photomicrographs of Hematoxylin & Eosin (H&E) stained cervical lymph node tissue section obtained from a patient;
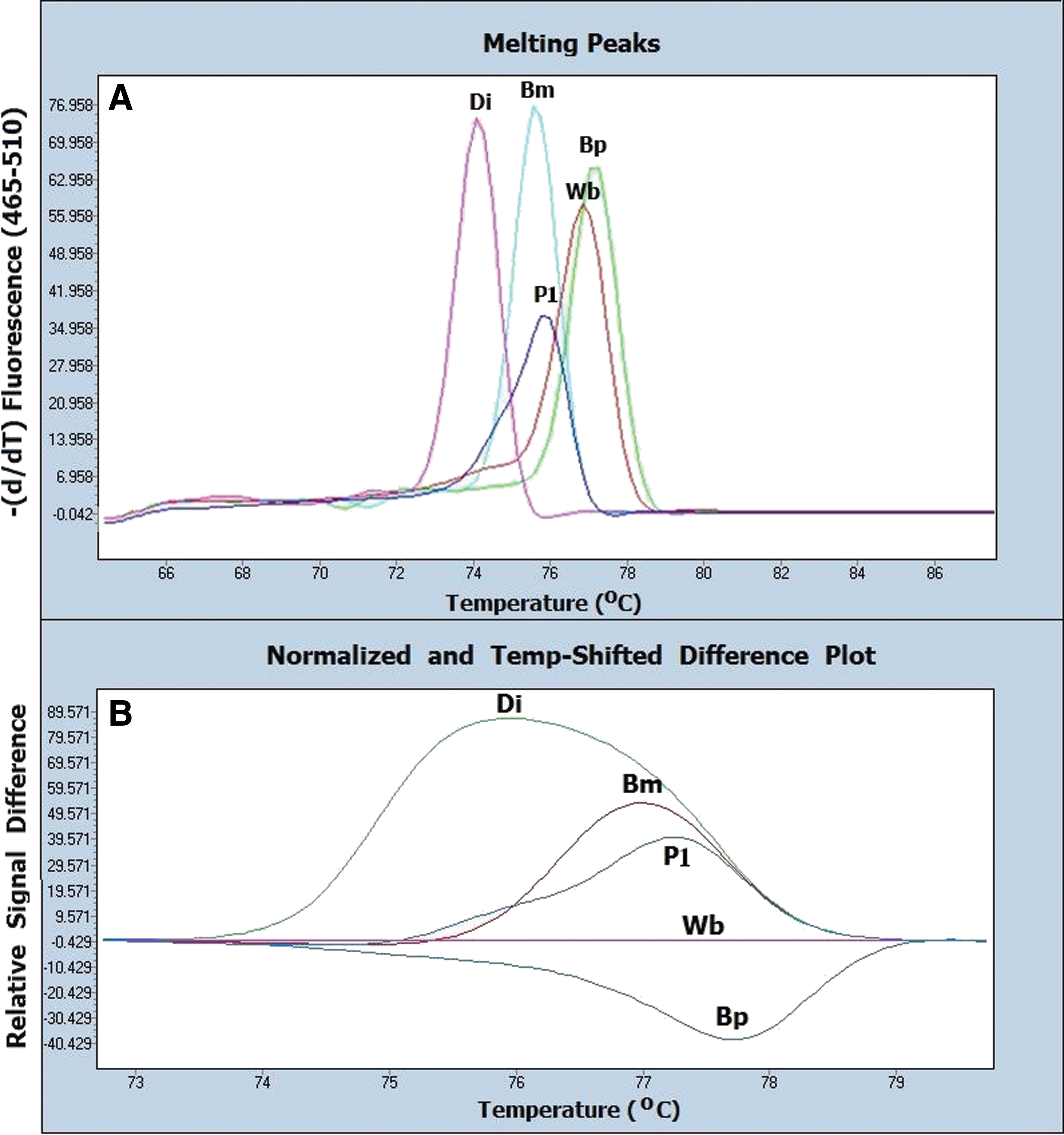
Results of the HRM assay for the diagnosis and identification of filarial infection in the unstained FFPES (DNA extraction using a quick paraffin-removal technique) and the H&E-stained FFPES are shown in Figure 4 and Table 1. The positive controls consisting of B. malayi, B. pahangi, D. immitis, and W. bancrofti were identified by HRM assay at different melting temperatures (Tm). Normalized and temperature-shifted difference plots classified the positive controls into five different groups. Melting peaks and difference curves derived from the normalization data of amplified products of filariae in the unstained and the H&E-stained FFPES were concordant with the parasite species. Melting peaks and difference curves derived from normalization data of amplified product of the H&E-stained tissue sections obtained from a patient are shown in Figure 5. DNA extracts from the unstained FFPES of other tissue dwelling parasites, i.e., A. cantonensis, third-stage larva of G. spinigerum, and C. cellulosae, showed negative results (Fig. 6). The corresponding results obtained are presented in Table 1.

Melting peaks and difference curves derived from the normalization data of amplified products of the positive controls (Di, D. immitis, Bm, B. malayi, Bp, B. pahangi, Wb, W. bancrofti), negative control (Neg) and studied samples. A1 and B1 reveal melting peaks and normalized and temperature-shifted difference plot of the unstained FFPES of the adult stages of D. immitis (qU1), B. malayi (qU2), and B. pahangi (qU3). A2 and B2 show melting peaks and normalized and temperature-shifted difference plot of the H&E stained FFPES of the adult stages of D. immitis (S1), B. malayi (S2), and B. pahangi (S3). The corresponding results are presented in Table 1. Color images available online at
Presents melting curve analyses
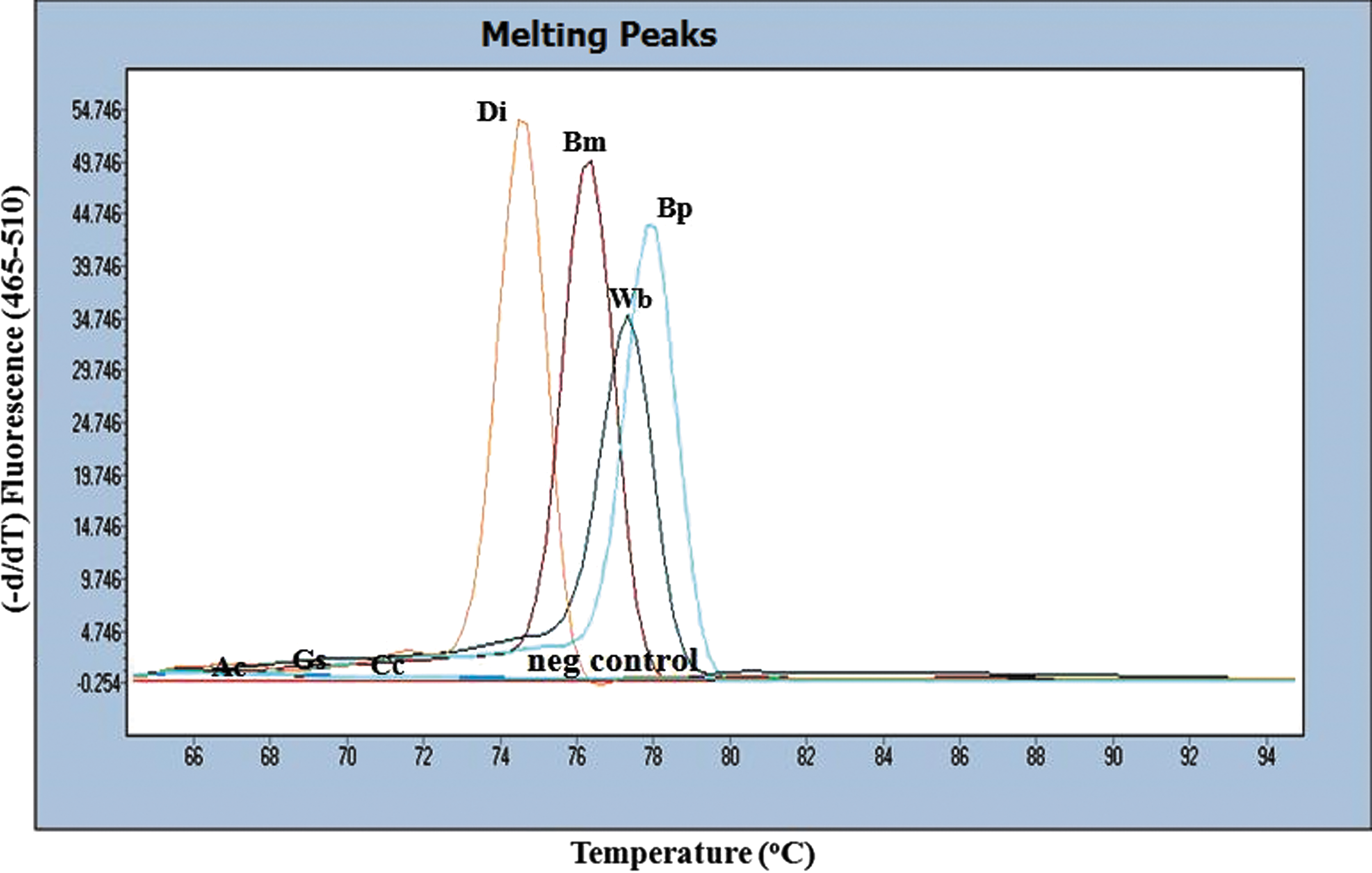
Melting peaks of the amplified products of the positive controls (Di, D. immitis, Bm, B. malayi, and Bp, B. pahangi); negative control (neg control); and samples of unstained FFPES of other tissue-dwelling parasites which showed negative results (Ac, A. cantonensis, Gs, G. spinigerum, Cc, C. cellulosae). The corresponding results are presented in Table 1. Color images available online at
H&E, Hematoxylin & Eosin; FFPES, formalin-fixed paraffin-embedded sections; Tm, melting temperature; SD, standard deviation; qU, amplified product of DNA extracted using the quick paraffin removal technique; sU, amplified product of DNA extracted using the standard paraffin removal technique.
Comparison of paraffin-removal methods
There was no difference in the use of the paraffin-removal or quick paraffin-removal protocol in the HRM-PCR assay of amplified products obtained from DNA extraction. Tm values of amplified products of unstained FFBES using DNA extract with a quick paraffin removal technique and unstained FFBES using DNA extract with a standard paraffin removal technique are shown in Table 1.
HRM-PCR specificity
The specificity of HRM-PCR was determined for all DNA extracts from the three other tissue-dwelling parasites. All samples were negative. No amplification curve was obtained (Table 1).
DNA sequencing
DNA sequences analyzed from the clinical specimen had 100% identity with B. malayi mitochondrial partial 12S rRNA gene obtained from GenBank (acc. no. AJ544843.1) and corresponded with HRM analysis.
Discussion
By using a real-time PCR with HRM analysis, both human and animal filarial parasites in FFPES were rapidly diagnosed and identified by Tm as well as normalized and temperature-shifted difference plots. Despite the close Tm values (difference <0.5°C) observed for W. bancrofti and B. pahangi, both species were discriminated using difference curves derived from the normalization data of amplified products using W. bancrofti as a baseline.
A molecular-based assay for the identification of other intact parasites in FFPE human tissues was recently reported using both conventional PCR and real-time PCR with probes (Koonmee et al. 2011, Rodriguez et al. 2014). The HRM-PCR assay provides a low-cost, fast, and sensitive molecular technique that allows the detection of DNA in a single step. The HRM reaction can be performed in a single tube (or well) using a single pair of primers, no specific probe is required and no post PCR is performed. The cost effectiveness of HRM is comparable to or cheaper than conventional PCR and sequencing (Dobrowolski et al. 2009; Zhou et al. 2005).
A previously designed primer pair used in the PCR amplified a 115-bp region of mitochondrial 12S rRNA gene from filarial worms. Subsequently, the HRM assay showed a specific Tm for each species. Previously, we reported the detection limitation of the assay was 1 microfilaria/reaction (Wongkamchai et al. 2013). The assay also had a high specificity because no false-positive results were detected in other FFPES prepared from other parasites.
In this study, an archived histopathological tissue containing structures of the female stage of a lymphatic filarial worm located in enlarged lymph node tissue located in the neck of a patient was used. The female filarial worm containing microfilariae within eggshells was identified only as an adult-stage lymphatic filaria because it lacked key characteristic morphology for species identification. By using the HRM-PCR assay, this case was identified as B. malayi. DNA sequencing of the patient sample showed 100% accordance with the HRM result, confirming that HRM is as effective as gene sequencing.
Most clinical laboratories use unstained sections for DNA extraction, whereas H&E-stained slides are only used as a reference. H&E staining allows visual confirmation of the presence of an organism in the section and reduces the risk of false-negative results. The possibility of using histochemically stained slides for DNA extraction has been explored by many studies and adverse consequences of the staining on DNA amplification have been reported. Morikawa et al. (2012) studied the DNA quality and assay performance of over 1000 H&E-stained FFPE cases and concluded that H&E staining had no effect on either DNA yield or molecular testing results. On the basis of their findings, in this study, H&E-stained sections were used for real-time PCR to identify filarial parasites. There was no difference in the Tm of amplified filarial DNA products from unstained FFPE or H&E-stained sections. Thus, our results are in concordance with Morikawa et al. (2012), because H&E staining did not affect the HRM-PCR assay results.
DNA extraction from FFPES is a critical step in a molecular assay. Potential problems, including paraffin as a physical barrier, DNA cross-linking, and PCR inhibitors, can affect the amplification of specific DNA fragments from specimens (An et al. 1991, Steinau et al. 2011). Paraffin removal is another obstacle during specimen processing for molecular testing. The standard protocol for paraffin removal requires a long time and is a multistep method. We developed a quick paraffin removal protocol that shortened the protocol length by about 30 min and had fewer steps compared with the standard protocol. When the quick paraffin removal protocol was compared with the standard protocol, no differences in the amount of DNA obtained or results of the real-time PCR assay were observed.
One disadvantage of H&E-stained sections is that tissues can flake off the side as a dry powder during scraping. Thus, a Pinpoint solution can be used to minimize potential contamination from tissue flakes in the work area (Snow et al. 2014). In the present study, we prevented flaking by adding lysis buffer directly onto the section prior to scraping.
The application of HRM real-time PCR for the diagnosis and identification of suspected filarial parasites in FFPES has the advantage of providing an effective diagnostic procedure for pathologists. Nevertheless, we acknowledge some limitations due to the limited availability of clinical specimens. To overcome this, we used adult filarial worms as well as other tissue-dwelling parasites in FFPES, which were prepared using the same method as for the clinical samples.
Further studies should apply the developed HRM-PCR assay for FFPES using greater numbers of clinical FFPE specimens. Moreover, the molecular-based diagnosis method used here would also be applicable for the identification of other parasites intact in organs or tissues.
The results obtained from the present study indicated that: (1) HRM-PCR is a useful method for identification and differentiation of filarial infection in FFPE specimens; (2) the assay was specific; (3) DNA was extracted with a quick paraffin removal technique and was comparable to the standard paraffin removal technique; and (4) H&E staining did not affect the HRM-PCR assay results.
Footnotes
Acknowledgments
This study was supported by grants from the Faculty of Medicine Siriraj Hospital, Mahidol University (grant no. R015732002). Thanks are also extended to Assoc. Prof. Dr. Boonyarit Cheunsuchon and Assoc. Prof. Dr. Piyavadee Leksrisakul, Department of Pathology, Siriraj Hospital, for kindly providing an archived histopathological tissue specimen. We also thank Miss Nuntiya Munkong and Miss Bungion Sermsart for laboratory assistance.
Author Disclosure Statement
No competing financial interests exist.