
Abstract
Although bats have been implicated as reservoir hosts for a number of zoonotic and life-threatening viruses, the bat bacterial flora and its zoonotic threat remain elusive. However, members of the vector-borne bacterial genera Bartonella causing various human as well as animal diseases have recently been isolated or detected from bats and their ectoparasites. In this study, we sampled 124 insectivorous microbats (Daubenton's bat, Myotis daubentonii) for peripheral blood in southwestern Finland in 2010. A Bartonella-specific PCR targeting rpoB (RNA polymerase β-subunit) was positive with blood samples from 46 bats (prevalence 37%). Scaled mass indexes of the infected and noninfected bats did not differ (p = 0.057). One rpoB sequence was identical with the rpoB sequence of B. naantaliensis strain 2574/1, previously isolated from bats in Finland. The rest of the sequences were highly similar to each other with nucleotide identity scores of 96% or higher. Nucleotide identity scores to the previously described type strain sequences of Bartonella or other database entries were no higher than 87%. Sequence analyses of another gene, gltA (citrate synthase), gave no higher than 90% nucleotide identity scores. On the basis of the conventional 95% sequence similarity cutoff in bacterial species delineation, a novel species of Bartonella was detected. We propose a species name Candidatus B. hemsundetiensis. Phylogenetic analyses based on rpoB and gltA sequences indicate that Candidatus B. hemsundetiensis clusters in a deep-branching position close to the ancestral species B. tamiae and B. bacilliformis. Our study reinforces the importance of bats as reservoirs of Bartonella.
Introduction
Bartonella
Although bats provide significant ecosystem services, such as pollination, seed dispersal, and pest control (Boyles et al. 2011), they have received a lot of negative attention due to their potential as reservoirs of zoonotic diseases. Research has mainly focused on viruses, such as severe acute respiratory syndrome (SARS) coronaviruses, Ebola and Marburg filoviruses, and Nipah and Hendra paramyxoviruses (Plowright et al. 2015). However, heterogeneous Bartonella strains from bats and their ectoparasites have recently been detected and isolated worldwide, e.g., (Bai et al. 2015). We have isolated and detected with PCR Bartonella strains from insectivorous microbats as well as their ectoparasites in Finland (Veikkolainen et al. 2014), which closely resemble a strain (Candidatus B. mayotimonensis) detected in a resected aortic valve tissue of an endocarditis patient in the United States (Lin et al. 2010). The present study aimed to analyze the prevalence of Bartonella colonization in the insectivorous Daubenton's bat subpopulations encountered in southwestern Finland.
Materials and Methods
Bat sampling and molecular detection of Bartonella
Bats were captured as described under animal ethics committee licence ESLH-YM-2007-01055 (Veikkolainen et al. 2014). Bats were identified visually to species and measured for mass and forearm length. These values were used to calculate the scaled mass index (SMI) (Peig and Green 2010), a useful numeric value to compare the overall health and nutritional status of the bats. Statistical analysis of SMI comparisons were performed with IBM SPSS Statistics version 22. A peripheral blood sample was collected into a 75-μL heparinized capillary tube from the interfemoral vein after lancing with a 25-gauge needle. All bats were released after sampling.
DNA samples were prepared from the blood samples stored at −80°C as described (Veikkolainen et al. 2014). An amount of 100–300 ng of DNA was used as templates in a Bartonella-specific master mix–based PCR targeting rpoB as described (Oksi et al. 2013). Part of the rpoB-positive samples were also subjected to a Bartonella-specific master mix–based PCR targeting gltA as described (Oksi et al. 2013). Distilled water and chromosomal DNA of B. henselae strain Houston 1 (Table S1; Supplementary Data are available at
Nucleotide sequence and phylogenetic analyses
Bartonella species included in the analyses are shown in Table S1. Sequences of the rpoB were trimmed down to 366-bp fragments (corresponds to nucleotide positions 246–610 of B. alsatica rpoB, AF165987) to match the length of the obtained rpoB sequences from the bat blood samples. Sequences of the gltA were trimmed down to 313-bp fragments (corresponds to nucleotide positions 4–315 of B. alsatica gltA, AF204273) to match the length of the gltA sequences obtained from the bat blood samples. Phylogenetic analyses were performed with concatenated rpoB and gltA sequences using Molecular Evolutionary Genetics Analysis (MEGA) 5.2.1 (available at
Results
A total of 124 Daubenton's bats (79 males and 45 females) were sampled in a geographical area of 2500 km2 in the archipelago of southwestern Finland in 2010 (Fig. 1, Table S2). Bartonella-specific PCR targeting rpoB was positive and was sequence verified with blood samples of 46 bats (prevalence 37%). In all, 38% and 36% of the males and females, respectively, were Bartonella positive. Bartonella-positive bats were detected in all of the 14 sampling sites. The mean SMI values of infected and noninfected bats were 8.7596 (standard deviation [SD] ±0.8858) and 8.3964 (SD ±1.0842), respectively, with an independent samples t-test p value of 0.057.
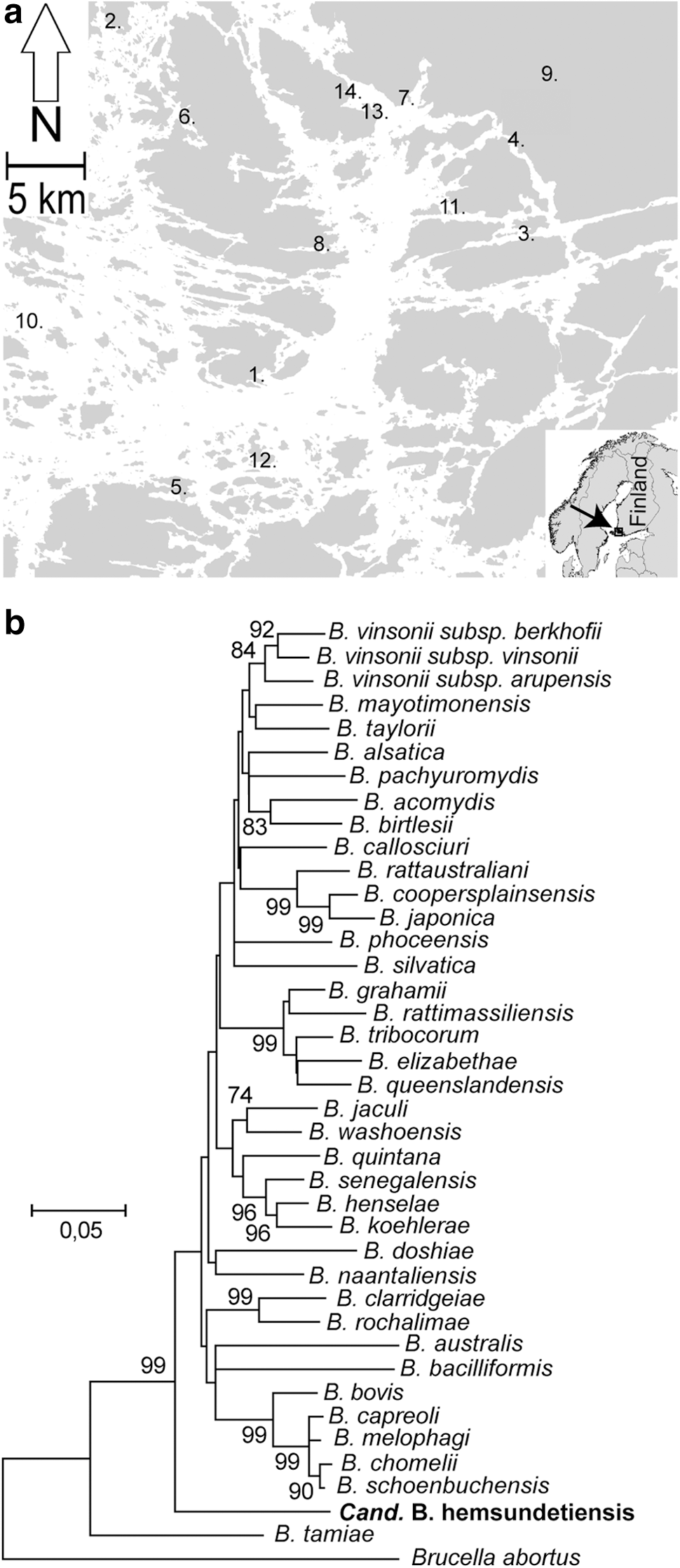
Study area and phylogenetic position of the Candidatus B. hemsundetiensis. (
Six unique rpoB alleles were detected (GenBank acc. nos. KP681546–KP681551). One rpoB allele was identical with the rpoB sequence of B. naantaliensis strain 2574/1, previously isolated from Daubenton's bat in the same geographical area (Veikkolainen et al. 2014). The rest of the sequences were highly similar to each other with nucleotide identity scores of 96% or higher. Pairwise genetic distance values of rpoB as compared to the Bartonella type strain sequences are shown in Table S3. On the basis of the conventional 95% sequence similarity cutoff in bacterial species delineation as proposed by Goris and co-workers (Goris et al. 2007), these Bartonella strains classify as strains of a single species. Most notably, nucleotide identity scores to the previously described type strain sequences of Bartonella or other database entries did not rise higher than 87%. This similarity percentage was detected, with the most prevalent (63%) rpoB allele KP681546 with an rpoB fragment (GU143508) amplified from a lung tissue of an Asian house shrew (Suncus murinus) in Nepal, Kathmandu. Sequence analyses of another gene, gltA (citrate synthase), gave no higher than 90% nucleotide identity scores. Pairwise genetic distance values of gltA as compared to the Bartonella type strain sequences are shown in Table S4. Phylogenetic analyses based on concatenated rpoB and gltA sequences indicated that the novel bat Bartonella clusters in a deep-branching position close to the ancestral species B. tamiae and B. bacilliformis (Fig. 1).
Discussion
We report that Daubenton's bats are a potent environmental reservoir of Bartonella in the archipelago of southwestern Finland (prevalence of 37%). Bartonella infection did not appear to affect negatively the overall health of the bats as analyzed by scaled mass indexing. Although we recently isolated several strains of Bartonella from bats in the same area in 2012, the majority of the Bartonella rpoB sequences detected in 2010 are distantly related (87% or less) to the Bartonella rpoB database sequences. Yet, all of these new rpoB sequences could be classified as coming from a single species, when the conventional arbitrary 95% rule of bacterial species delineation was applied (Goris et al. 2007). Therefore, we propose that majority of the rpoB sequences were originating from a novel species of Bartonella Candidatus B. hemsundetiensis. The name is based on one bat sampling location of this study, Hemsundet (see Fig. 1). Our attempts to isolate strains of this new species failed using the conventional blood agar method previously successful in our bat project (Veikkolainen et al. 2014). Isolated strains would allow detailed multilocus sequence analyses, and therefore more reliable analyses of phylogenic relationships to other members of the genus Bartonella. The role of ectoparasites in the maintenance and transmission of Candidatus B. hemsundetiensis colonization in Daubenton's bat and other bat species should be explored in future.
Footnotes
Acknowledgments
We thank all the students and volunteers at the University of Turku who helped collect the bat peripheral blood samples.
Author Disclosure statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.