
Abstract
Plague is a disease of epidemic potential that may emerge with discontinuous outbreaks. In South America, 50 wild rodent species have been identified as plague reservoirs, in addition to one lagomorph and two marsupials. To review the nomenclature of plague reservoirs, we examined specimens collected in plague foci, carried out new surveys in Brazilian plague regions, and re-evaluated the nomenclature of South American reservoirs on the basis of the current literature. Five of the 15 species involved with plague in Argentina, three of 10 species involved with plague in Bolivia, three of the seven species involved with plague in Peru, five of the nine species involved with plague in Ecuador, and six of the nine species involved with plague in Brazil have undergone taxonomic changes. In the last 20 years, plague cases were recorded in Bolivia, Brazil, Ecuador, and Peru. These four countries have a high rodent species richness in plague foci, a fact that may be decisive for the maintenance of plague in the wild.
Introduction
P
Plague was introduced in South America through seaports during third pandemic that started in China during the 19th century first in Paraguay, in 1899, and persisted for more than 50 years in Argentina, Venezuela, Bolivia, Brazil, Ecuador, and Peru (Pollitzer 1954, Schneider et al. 2014). The diversity of rodent fauna and their parasitic fleas has been responsible for plague fluctuations in rodent reservoirs and humans, pointing to the relevance of a permanent assessment of the rodent fauna in afflicted countries (Stenseth et al. 2008).
Yersinia pestis is a serious threat, still affecting thousands of humans every year. The effect of climate change on plague transmission has been well studied and potentially induces increased incidence in natural reservoirs (Stenseth et al. 2006, 2008), geographical range shifts, or reductions in the incidence (Snall et al. 2009). The persistence of enzootic cycles within relatively small rodent populations is one of the main factors explaining the historical persistence of plague, despite long disease-free periods (Keeling and Gilligan 2000). Plague cannot be eradicated because its agent survives in fleas and soils and is mostly widespread in wild reservoirs; thus, understanding the dynamics of these reservoirs is crucial (Parmenter et al. 1999, Enscore et al. 2002).
Serologic and/or bacteriological evidence of Y. pestis infection in rodents has been reported in Ecuador, Peru, and Brazil (Ruiz 2001, Tavares et al. 2012). Because the taxonomy of many small mammal species has been revised, new taxonomic arrangements have been proposed, and new species and genera have been described. The identification of plague reservoirs and the association of recent records to previously published information has been a confusing task. Thus, it is of paramount importance to continuously revise and update the taxonomic status of rodent reservoirs. To update the nomenclature of South American plague reservoirs, we collected small mammals in plague-afflicted regions of Brazil and examined specimens of the mammal collection of Museu Nacional (MN), Universidade Federal do Rio de Janeiro, which had been obtained in plague foci areas (Oliveira and Franco 2005).
Materials and Methods
Specimens were collected in Brazilian plague foci (Fig. 1), and the identification, on the basis of morphology, was confirmed by karyotype for selected specimens following the protocol of Andrade and Bonvicino (2003). Voucher specimens were deposited in the mammal collection of the MN and in the Laboratório de Biologia e Parasitologia de Mamíferos Reservatórios Silvestres, IOC/Fiocruz.

Map with the geographic distribution of the main rodent plague host species in South America. Shaded areas are human plague foci adapted from Aragão et al. (2007) and collecting sites in Brazilian plague foci.
Select specimens deposited in the MN were revised, and an updating of the taxonomy of plague reservoirs from South America was carried out. Evidence of the species as plague reservoirs was assessed by using official information from the World Health Organization, the Pan American Health Organization (PAHO), Ministries of Health, and others scientific publications.
Results and Discussion
Plague-associated rodents and lagomorphs in Argentina
Argentina was afflicted by plague in 1899 after infection in the Paraná River ports that subsequently spread inland (Sussini 1939, World Health Organization 1965). At least 14 rodent species were found to be plague positive (Table 1, Figs. 1 and 2), with Graomys centralis (= Graomys chacoensis) playing a relevant role in spreading the infection among rodents (de la Barrera 1953). Calomys specimens previously identified as Hesperomys, a former genus that included species currently allocated to 21 genera, were plague-positive in Argentina (World Health Organization 1959): H. bimaculatus (= C. laucha), H. laucha (= C. laucha), H. murillus cordovensis (= C. musculinus) and H. venustus (C. venustus). Other sigmodontines include Holochilus balnearum (= H. chacarius), Oryzomys flavescens (= Oligoryzomys flavescens), Akodon dolores, Eligmodontia moreni, and Graomys griseoflavus. Histricognath rodent species were also involved, including Lagostomus maximus, Cavia aperea, Microcavia australis, and Galea musteloides (Table 1), as well as the lagomorph Sylvilagus brasiliensis, which was rarely found to be infected (de la Barrera 1953).
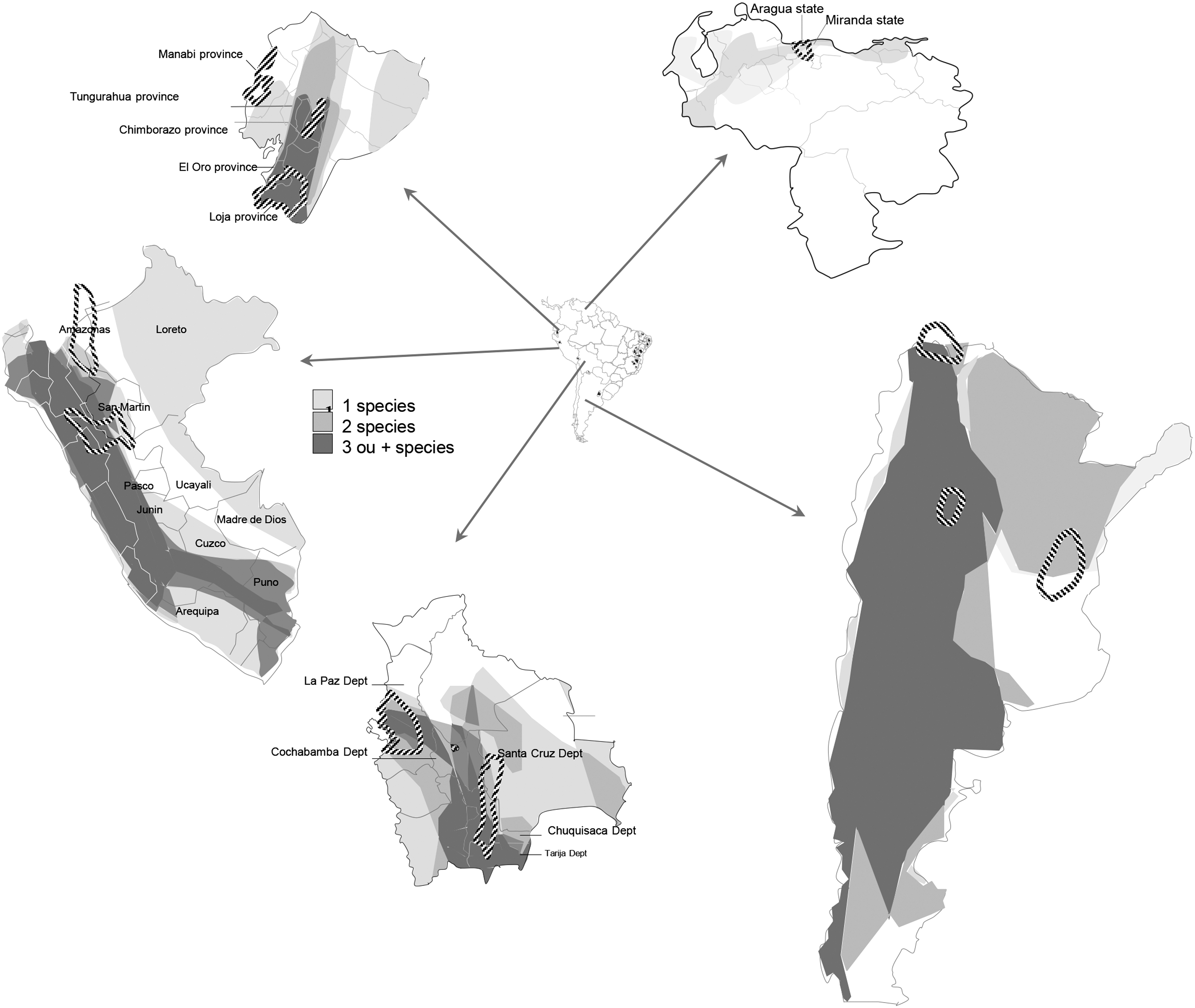
Map with the geographic distribution of the main rodent plague host species in Argentina, Bolivia, Ecuador, Peru, and Venezuela.
Pos/Tot, number of infected individuals over the total number captured.
Argentinean (AR) provinces of Buenos Aires (BA), Córdoba (CD), La Pampa (LP), Mendoza (MZ), Río Negro (RN), Sanla (SA), Santiago del Estero (SE); Bolivia (BO); Ecuadorian (EC) provinces of Chimborazo (CB) and Loja (LJ); Peruvian (PE) department of Píura (PIU); Venezuelan (VE) states of Aragua (AR), Miranda (MI); and Brazilian (BRA) states of Alagoas (AL), Ceará (CE), Paraíba (PB), and Pernambuco (PE).
1 = Jervis-Alarcón (1958), 2 = Macchiavello (1957), 3 = World Health Organization (1965), 4 = World Health Organization (1959), 5 = Freitas (1970), 6 = Almeida et al. (1987), 7 = de la Barrera (1953), 8 = Machiavello (1959), 10 = Almeida et al. (1989), 11 = Martin-Alonso et al. (2014), 12 = Almeida et al. (1984).
Correspondence between number of animals and locality of collection.
Plague-associated rodents and lagomorphs in Bolivia
Bolivia was afflicted by plague in 1921, probably conveyed by rodents from Argentina, subsequently spreading to the north (World Health Organization 1965). At least 10 small mammal species were involved with plague in Bolivia (Table 1; Figs. 1 and 2). Graomys griseoflavus and Oxymycterus paramensis retained this taxonomy. Some were assigned to other genera: Calomys fecundus (= Calomys boliviae) was previously assigned to Hesperomys and O. flavescens to Oryzomys. C. fecundus was later considered a junior synonym of C. venustus or C. bolivae (Musser and Carleton 2005) or a valid species (Dragoo et al. 2003, Almeida et al. 2007, Patton et al. 2015). Calomys venustus is presently considered endemic to Argentina (Patton et al. 2015), and specimens identified as C. venustus in Bolivia are likely to be C. boliviae.
Oligoryzomys flavescens (World Health Organization 1959; Table 1), reported as plague-positive in Bolivia (World Health Organization 1959), is restricted to Uruguay, Paraguay, Argentina, and Brazil (Fig. 1), ruling out that this species was the plague reservoir in Bolivia. S. brasiliensis is also plague positive in Bolivia.
Rodents and lagomorphs involved in plague in Peru
Plague arrived in Peru in 1903 through seaports and spread inland (Macchiavello 1957, World Health Organization 1965). Widespread epidemics have occurred recently, including primary pneumonic plague with nosocomial transmission (Dávalos et al. 2001, Donaires et al. 2010, Martin-Alonso et al. 2014).
At least seven rodents and one lagomorph (S. brasiliensis) species were plague-positive in Peru (Table 1, Figs. 1 and 2). Sigmodon peruanus, and the more abundant Akodon mollis and Oryzomys xantheolus (= Aegialomys xantheolus), were frequently found infected (World Health Organization 1965). Species involved in plague events were assigned to new genera, such as A. xantheolus, Oligoryzomys andinus, and Oligoryzomys arenalis previously included in the genus Oryzomys. Rhipidomys equatoris is currently regarded as a synonym of Rhipidomys leucodactylus, and Akodon mollis orophilus has been given full specific rank as Akodon orophilus (Table 1). Recently, Hylaeamys perenensis and Oecomys spp. were found plague-positive in Peru (Martin-Alonso et al. 2014).
Rodents and lagomorphs associated with plague in Ecuador
Plague was recorded in Guayaquil in 1908, spreading inland between 1909 and 1939 (Jervis-Alarcón 1958, 1967). The cultural habits of killing fleas with teeth bites and raising guinea pigs inside homes, in addition to incidental consumption of infected guinea pigs, all contribute to plague epidemics among the Andean population (Gabastou et al. 2000, Ruiz 2001, Schneider et al. 2014).
A. mollis and A. xantheolus are plague resistant and the most abundant hosts of plague in Ecuador (World Health Organization 1965). Sciurus stramineus (= Simosciurus straminaeus), another plague-resistant species, plays an important role as host in Ecuador, mainly in the Loja Province, whereas Sylvilagus brasiliensis, a comparatively resistant species, is the main reservoir in Chimboraza Province (World Health Organization 1965). A. mollis has had a more stable nomenclature, but most plague-involved species have been assigned to different genera, such as Aegialomys xantheolus, Oligoryzomys longicaudatus, and Oligoryzomys flavescens, all previously included in Oryzomys. The plague host Phyllotis fruticicolus is currently referred as Phyllotis andium, whereas Sigmodon puna is currently allocated to Sigmodon peruanus (Table 1, Figs. 1 and 2).
Rodents involved in plague in Venezuela
Plague appeared in 1908 in Venezuela, in the port of La Guaira, spread to Caracas in 1919, reaching rural areas in 1910 and adjacent regions of Aragua state in 1939, with continuous spurts from time to time until 1951. Plague foci were found in the states of Aragua and Miranda, where Sigmodon hirsutus and Heteromys anomalus (Figs. 1 and 2) were the reservoirs (World Health Organization 1959, 1965).
Rodents and marsupials involved in plague in Brazil
Plague was introduced in Brazil by sea route in 1899, becoming endemic in rural areas and affecting human populations until the mid-1980s, when all foci showed a tendency to quiescence (World Health Organization 1965, Tavares et al. 2012). The latest significant events of human plague occurred in the Brazilian states of Ceará and Paraíba in the 1980s. In the 1990s and in 2005, human cases were confirmed in Ceará; Bahia State reported human plague cases until 2000 (Tavares et al. 2012).
Rodents were monitored for plague for several years, with many species identified as reservoirs or hosts (Table 1). The main host species in the Brazilian northeastern foci is Necromys lasiurus (previously included in Akodon, Bolomys, or Zygodontomys, and which currently includes Zygodontomys pixuna as a junior synonym; Table 1, Fig. 1).
Thrichomys apereoides (Table 1) was previously referred to as Cercomys cunicularius until Petter (1973) invalidated this name. The genus Thrichomys was considered monospecific for a long time despite the high number of subspecies recognized. Recently, the remarkable genetic diversity in Thrichomys led to the recognition of the former subspecies as full species (Nascimento et al. 2013) (Fig. 1). Thrichomys populations from Ceará, Piauí, Paraíba, and Pernambuco belong to T. laurentius, which is a plague reservoir in these states (Fig. 1). Specimens identified as Cercomys inermis (= Thrichomys inermis) were found infected in Bahia (World Health Organization 1959). Because T. laurentius also occurs in Bahia, further studies are necessary to identify which species of Thrichomys is the plague reservoir in this Brazilian state.
Specimens identified as Oryzomys subflavus were found infected with plague in Pernambuco and Paraíba (Table 1). Neotropical species previously assigned to the genus Oryzomys have been sorted into 10 new genera in the last comprehensive review of Oryzomyini. The genus Cerradomys was erected to accommodate the Oryzomys subflavus species-group (Weksler et al. 2006). Morphologic and genetic studies of this species-group led to the recognition of seven new species. Cerradomys langguthi is the plague reservoir in northeastern Brazil (Fig. 1), and the most common rodent genus collected in sampled areas (Table 2).
The uppermost line indicates the region followed by the state in parentheses, followed by municipalities.
Karyotyped specimen; n = sample size.
Calomys is a widespread genus with several species involved in plague enzootic cycles (Table 1). This was the case of specimens identified as Calomys callosus in Ceará and Pernambuco (Almeida et al. 1987), a species long considered the single large-sized Calomys occurring in Brazil. On the basis of morphology, Hershkovitz (1962) considered C. expulsus as a junior synonym of C. callosus; however, morphometric and karyotypic differences showed that they belong to different species (Bonvicino and Almeida 2000). In Brazil other Calomys species also occur (Fig. 1), C. expulsus being the only known plague reservoir (Fig. 1).
Oligoryzomys, previously considered a subgenus of Oryzomys, is distributed throughout a large portion of the Neotropics, and includes several species that are plague hosts in South American countries (Table 1). Three Oligoryzomys species occur in northeastern Brazil: O. nigripes, O. stramineus (Bonvicino and Weksler 1998), and O. mattogrossae J.A. Allen 1916 (Fig. 1). Whereas O. mattogrossae can be easily distinguished by morphology, the other species are very similar, mainly in fur coloration, sharing the whitish ventral pelage clearly delimited from the dorsal coloration, a character also shared with Oligoryzomys eliurus, a junior synonym of O. nigripes (Weksler and Bonvicino 2005). Specimens from southeastern Brazil, formerly identified as Oryzomys eliurus, were also plague positive (Table 1). O. nigripes is the single species with a whitish ventral pelage occurring in the area and is the plague reservoir in southeastern Brazil. Conversely, the identification of plague reservoirs in the northeastern region require further studies because specimens previously identified as O. eliurus may belong to either O. nigripes and O. stramineus, two morphologically very similar species (Fig. 1).
Plague-positive specimens referred originally to Zygodontomys lasiurus and Zygodontomys pixuna are currently assigned to Necromys lasiurus, a single species that constitutes an important plague reservoir in northeastern Brazil (Oliveira 1992) (Fig. 1). Our survey of plague foci in northeastern Brazil showed this species to be abundant (Table 2). Necromys lasiurus has had a confused taxonomic history, being assigned just in the last century to the genera Akodon, Zygodontomys, Bolomys, and Cabreramys, the last two currently sunk in the synonymy of Necromys.
Akodon specimens were found positive for plague in several South American countries (Table 1). Akodon cursor Winge 1887, an endemic Brazilian species (Bonvicino et al. 2008), occurs in the plague foci of Pernambuco, Paraíba, and Minas Gerais States. However, in Rio de Janeiro, several Akodon species are sympatric, including Akodon montensis Thomas 1913, and further studies are necessary to identify whether A. montensis, A. cursor, or both species might be the plague host in southeastern Brazil.
In northeastern Brazil, the best-studied region in terms of plague, there are two plague-resistant species—the commensal rat Rattus rattus and the wild guinea pig Galea spixii. These species may form the basis of enzootic foci, whereas the most sensitive rodent to Y. pestis is N. lasiurus, which is the amplifying host and the main species involved in large outbreaks (Karimi et al. 1974, 1976, Poland and Barnes 1979). Other Sigmodontinae species like Calomys expulsus, Holochilus sciureus, O. nigripes, and Cerradomys langguthi are also sensitive to Y. pestis, but few of these rodents have been detected infected (Almeida et al. 1987, 1984). N. lasiurus is scansorial, gramnivorous, a habitat generalist, and very common and abundant over all the Caatinga and Cerrado biomes (Bonvicino et al. 2008). N. lasiurus can reach high densities in some periods, and it is also an inhabitant of peridomestic and rural areas, increasing the possibility of human contact and, consequently, allowing the plague transmission to humans by its fleas. G. spixii is an herbivorous, terrestrial, common habitat specialist, and can be locally abundant, occurring in open grasslands, with or without rocky outcrops (Bonvicino et al. 2008).
Final Considerations
Despite outbreaks recorded at the beginning of the 20th century, plague apparently disappeared from Argentina, Chile, Paraguay, Uruguay, and Venezuela. In the last 20 years, plague cases have been recorded in Bolivia, Brazil, Ecuador, and Peru. Despite declining incidence, morbidity, and mortality in human populations, residual plague activity persists among host reservoirs and the possibility of recurrence cannot be discarded. Protection of human populations from a plague re-emergence requires maintenance of a continuous and systematic monitoring of rodent reservoirs. Current knowledge about the factors playing a role in the epidemiological chain events in South American countries is still unsatisfactory. Persistent plague activity in these areas should be considered a real and permanent threat to population health, which may extend to other regions, including urban centers. Ecological or social changes may trigger the recurrence of plague at any time.
We hope that this report will promote the continuation of studies aimed at clarifying the introduction, spread, establishment and permanence of plague in South American countries.
Footnotes
Acknowledgments
We are grateful to the staff of the secretary of health surveillance of Caruaru, Exu, and Fortaleza for facilities. H. Seuánez provided insightful comments on the manuscript and improved the language style and grammar. Permits for field collection were granted by SISBIO under number 11375-1. This work is supported through grants from Conselho Nacional de Pesquisa e Desenvolvimento (processes numbers 303422/2010-6; 481286/2011-0; 308271/2013-0), Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro for CRB (E-26/110.505/2012, E-26/101.950/2012), and SVS/MS TC210/2011 process number 24382.000448/2012-59 for AMPA.
Author Disclosure Statement
No competing financial interests exist.