
Abstract
Culex (Cx.) pipiens mosquitoes are important vectors of West Nile virus (WNV). In Europe, the species Cx. pipiens consists of two biotypes, pipiens and molestus, which are morphologically identical, but differ in behavior. Typical behavior of the molestus biotype is the ability to remain active during winter, whereas the pipiens biotype enters diapause. The current paradigm is that the two biotypes occur sympatrically in southern Europe, but occur in isolated above- and belowground populations in northern Europe. In northern Europe, hybridization between biotypes is considered to be low because of the barrier that exists between typical habitats. Data on the occurrence of the biotypes and hybrids in northern Europe, however, are scarce, because identification to the level of biotype is often not performed. Our objective was to clarify the distribution of the Cx. pipiens biotypes and to determine hybridization rates in The Netherlands. Cx. pipiens mosquitoes were collected using three different approaches. First, traps were deployed randomly throughout The Netherlands during the summers of 2011 and 2012 (active surveillance). Second, using a web-based reporting platform and media campaign, Dutch citizens were asked to send dead mosquitoes to our laboratory during the winter and summer of 2014 (passive surveillance). Third, larvae and adults were collected during the summer of 2014 from aboveground locations in Amsterdam to identify molestus larval habitats. Real-time PCR was used for identification to the level of biotype. We found that biotype molestus and hybrids were feeding indoors during winter and summer in The Netherlands and that hybridization rates ranged between 6% and 15%. Larval habitats of biotype molestus were found to occur aboveground. The high percentage of hybridization has implications for assessing the risk of WNV transmission, because hybrids are thought to have ideal characteristics for bridging WNV between birds and humans.
Introduction
W
Culex (Cx.) pipiens mosquitoes have been identified as important vectors of WNV in the United States (Fonseca et al. 2004, Kilpatrick et al. 2005). The species, Cx. pipiens, consists of two biotypes named pipiens (Linnaeus 1758) and molestus (Forskål 1775), which are identical in morphology but have different genetic backgrounds and show distinct behavior. The pipiens biotype occurs aboveground, prefers birds as hosts (ornithophilic), mates in swarms (eurygamous), requires a blood meal to lay eggs (anautogenous), and diapauses during winter (Byrne and Nichols 1999). In contrast, the molestus biotype occurs belowground (e.g., in subway tunnels), prefers mammals as hosts (mammophilic), mates in confined spaces (stenogamous), does not require a blood meal to lay the first egg batch (autogenous), and remains active during winter (Marshall and Staley 1935, 1936, Shute 1941, Willcox 1941). Host availability also plays an important role in the selection for hosts. Approximately 90% of females of the mammophilic molestus biotype that were collected from inside chicken coops in Portugal had blood-fed on birds (Gomes et al. 2013). Hybrids between the pipiens and molestus biotypes are hypothesized to have intermediate behavioral traits, and this may favor the transmission of WNV from birds to humans (Fonseca et al. 2004, Kilpatrick et al. 2005). Hybridization, however, is considered to be low in northern Europe due to separation of the typical habitats of the two biotypes (Fonseca et al. 2004). If hybrids indeed play a key role in WNV transmission, then the absence of hybrids could be one of the possible explanations for the absence of WNV cases in northern Europe.
Although several studies have measured the spatial distribution of Cx. pipiens and the related species Cx. torrentium (Martini 1925) in Europe, most of them did not differentiate to the biotype level (Hesson et al. 2011, Weitzel et al. 2011, Hesson et al. 2013). The view of spatially separated populations of the two biotypes (Fonseca et al. 2004), therefore, may not reflect the real situation in northern Europe, because differentiation between biotypes was not addressed in many studies. Rudolf et al. (2013) performed analyses to the biotype level and found that the molestus and pipiens biotypes occur sympatrically in Germany. No strong conclusions on the occurrence of biotype molestus or hybrid frequencies could be drawn, however, because samples were analyzed in pools and habitats were not systematically analyzed (Rudolf et al. 2013).
The main objective of the current study was to elucidate the ecology of the biotypes of Cx. pipiens and their hybrids in northern Europe, with a focus on the spatial and seasonal distribution in The Netherlands. Developing our understanding of the occurrence of the different Cx. pipiens biotypes and hybrids is essential for WNV risk assessments because not all mosquitoes are expected to be equally competent vectors or have equal inherent characteristics for bridging WNV to humans.
Materials and Methods
Adult mosquito collection: Active surveillance throughout The Netherlands
The Centre for Monitoring of Vectors operates an annual nationwide mosquito surveillance program in The Netherlands. The detailed protocol and outcomes of this program have been published elsewhere (Ibañez-Justicia et al. 2015). In brief, mosquitoes were sampled from mid-April until the second half of October in both 2011 and 2012, using CO2-baited Mosquito Magnet Liberty Plus traps (Woodstream Corp., Lititz, PA; hereafter MM-trap) for 375 randomly selected locations in The Netherlands (Ibañez-Justicia et al. 2015). At each location, one trap was run for 7 days, and thereafter the contents of the traps were collected and sent to the laboratory. Mosquitoes were identified morphologically on the basis of the Culicidae key specifically designed for rapid field identification of Dutch adult Culicidae (modified after Snow 1990, Schaffner et al. 2001, Verdonschot 2002, Becker et al. 2010). Mosquitoes were caught at 353 locations (94.1%), of which in total 3363 Cx. pipiens/torrentium mosquitoes were caught at 236 (62.9%) of these locations. At least one Cx. pipiens/torrentium sample from each of 158 locations was available and in good condition for further molecular analysis. One mosquito of each of these locations (2011, n = 82; 2012, n = 76) was, therefore, selected for further analysis to the species and biotype level by real-time PCR (see below).
Adult mosquito collection: Passive surveillance throughout The Netherlands
A different and unique sampling method was developed for indoor mosquito collections, involving the participation of Dutch citizens (Kampen et al. 2015). A website was launched through which Dutch citizens were asked to fill out a questionnaire regarding mosquito presence and possible nuisance (
A selection of 150 Cx. pipiens/torrentium samples, divided over two groups, was made of the samples that were sent in by citizens during the winter of 2014. Mosquitoes in the first group (n = 68) had caused self-reported nuisance and by visual inspection upon arrival in the laboratory were confirmed to be blood-fed (active during winter), whereas mosquitoes in the second group (n = 82) had not caused nuisance and were not engorged. For the samples submitted during summer, a selection based on the same criteria, in total 172 Cx. pipiens/torrentium samples, was made. Mosquitoes in the first group (n = 89) had caused nuisance and by visual inspection were confirmed to be blood-fed, whereas mosquitoes in the second group (n = 83) had not caused nuisance and were not engorged. All selected mosquitoes were further analyzed by real-time PCR (see below).
Larval and adult mosquito collection: In-depth study of the city of Amsterdam
The highest number of mosquito reports came from Amsterdam and this triggered further investigation. A stratified random sampling design (1 × 1 km) was used to select 94 locations in Amsterdam. From May to September of 2014, all publicly accessible larval habitats were identified within a radius of 50 meters around each selected location. This resulted in 133 habitats where mosquito larvae were found. Fourth instar larvae were identified to the species level on the basis of morphology following Becker et al. (2010). In total, 119 larval habitats contained Cx. pipiens larvae. From a selected number of 48 habitats, all larvae (n = 480) were identified to the biotype level by real-time PCR (see below). In addition, from July to September of 2014, MM-traps were placed at a selection of 20 out of the 94 locations for a duration of 2 weeks. All Cx. pipiens/torrentium adults captured in these traps (n = 161) were also identified to the biotype level by real-time PCR.
Molecular identifications: DNA extraction
DNA was extracted by heating mosquito legs in 100 μL of 4 M ammoniumhydroxide for 20 min at 99°C, cooling on ice for 1 min, centrifuging (20 s at 3700 rpm) and heating again for another 20 min at 90°C with the lids open (Guy and Stanek 1991). DNA extracts were stored in the freezer at −20°C until further analysis.
Molecular identifications: Primer and probe design
The real-time PCR assay as described by Rudolf et al. (2013) was adjusted to optimize the design, because amplification curves did not all show an exponential increase and did not reach the same plateau, which made the interpretation of results difficult. DNA sequences of microsatellite CQ11 of Cx. pipiens biotypes pipiens, molestus, pallens, and quinquefasciatus were mined from GenBank and aligned with Geneious R7 (Biomatters, New Zealand) using MAFFT (Katoh et al. 2002). On the basis of the alignments, new primers were designed with the use of Primer Express V3.0 software (Life technologies, The Netherlands). Cx_pip_F (5′-GCGGCCAAATATTGAGACTTTC-3′) and Cx_pip_R (5′-ACTCGTCCTCAAACATCCAGACATA-3′) were designed as universal Cx. pipiens forward and reverse primers, respectively. For identification of biotype molestus, probe Cpp_mol_P (5′-FAM-TGAACCCTCCAGTAAGGTA-MGB-3′) was designed, which is a shorter version of the original probe by Rudolf et al. (2013), conjugated with a minor groove binder (MGB) group. The two probes Cpp_pip_P1 (5′-VIC-CACACAAAYCTTCACCGAA-MGB-3′) and Cpp_pip_P2 (5′-VIC-ACACAAACCTTCATCGAA-MGB-3′) were designed for identification of biotype pipiens to cover the sequence variation at the original probe binding site. Hybrids can be identified by the presence of amplification curves for both the pipiens and molestus probes.
DNA sequences of acetylcholinesterase 2 (ace-2) from Culex spp. were mined from GenBank and aligned for Cx. torrentium identification. On the basis of the original primers and probes of Rudolf et al. (2013), forward and reverse primers were enlarged at the 5′ end to obtain Cx_tor_F (5′-CTTATTAGTATGACACAGGACGACAGAAA-3′) and Cx_tor_R (5′-GCATAAACGCCTACGCAACTACTAA-3′). The probe was shortened at the 5′ end and conjugated with a MGB group to get Cx_tor_P (5′-FAM-ATGATGCCTGTGCTACCA-MGB-3′).
Molecular identifications: Real-time PCR
Real-time PCR tests were performed in two separate reactions (one for each locus) and consisted of 1× Premix Ex Taq (Takara Bio, Japan), 300 nM of each primer, and 100 nM dual-labeled hydrolysis probe, except for the Cpp_pip1 for which 200 nM was added. Molecular grade water and 2 μL DNA template were added to reach a final volume of 25 μL. Thermocycler conditions were 95°C for 10 min, followed by 45 cycles of 95°C for 15 s and 62°C for 1 min. Real-time PCR runs were performed in 96-well plates on a CFX96 Real-Time PCR system (Bio-Rad Laboratories, Hercules, CA) and data were analyzed in CFX manager 2.0. Samples with Cq values below 40 and with real-time PCR curves showing exponential increase were considered positive. Positive amplification controls for each target organism and negative amplification controls were included in each real-time PCR run.
Molecular identifications: Validation
The new real-time PCR design was validated with at least 10 specimens from each line of morphologically and behaviorally well-characterized Cx. torrentium (Best, The Netherlands), biotype pipiens (Pennsylvania), biotype molestus (Bologna, Italy), and lab-reared hybrids between the latter two.
Distribution maps
Arc-GIS 10.0 (ESRI 2011; ArcGIS Desktop, Release 10, Redlands, CA, Environmental Systems Research Institute) was used to generate maps.
Results
Molecular identifications: Validation
The real-time PCR was initially performed with an elongation temperature (Ta) of 60°C. The obtained amplification curves for the characterized samples all showed exponential increases that matched the corresponding species or biotype and reached more or less the same plateau for each fluorophore. Cx. torrentium, however, gave a false positive signal with the biotype pipiens probes at high Ct (>39) values, although not with an exponential curve. Therefore a real-time PCR with a gradient (Ta = 60–63°C) was performed which had an optimum Ta of 62°C at which the false amplification curve for Cx. torrentium remained below the automatic threshold, whereas other curves were not affected.
Adult mosquito collection: Active surveillance throughout The Netherlands
Of the total number of 158 analyzed samples from the National Vector Survey, 104 samples were identified as biotype pipiens (65.8%), 39 as biotype molestus (24.7%), 10 as hybrids (6.3%), and five as Cx. torrentium (3.2%) (Fig. 1).
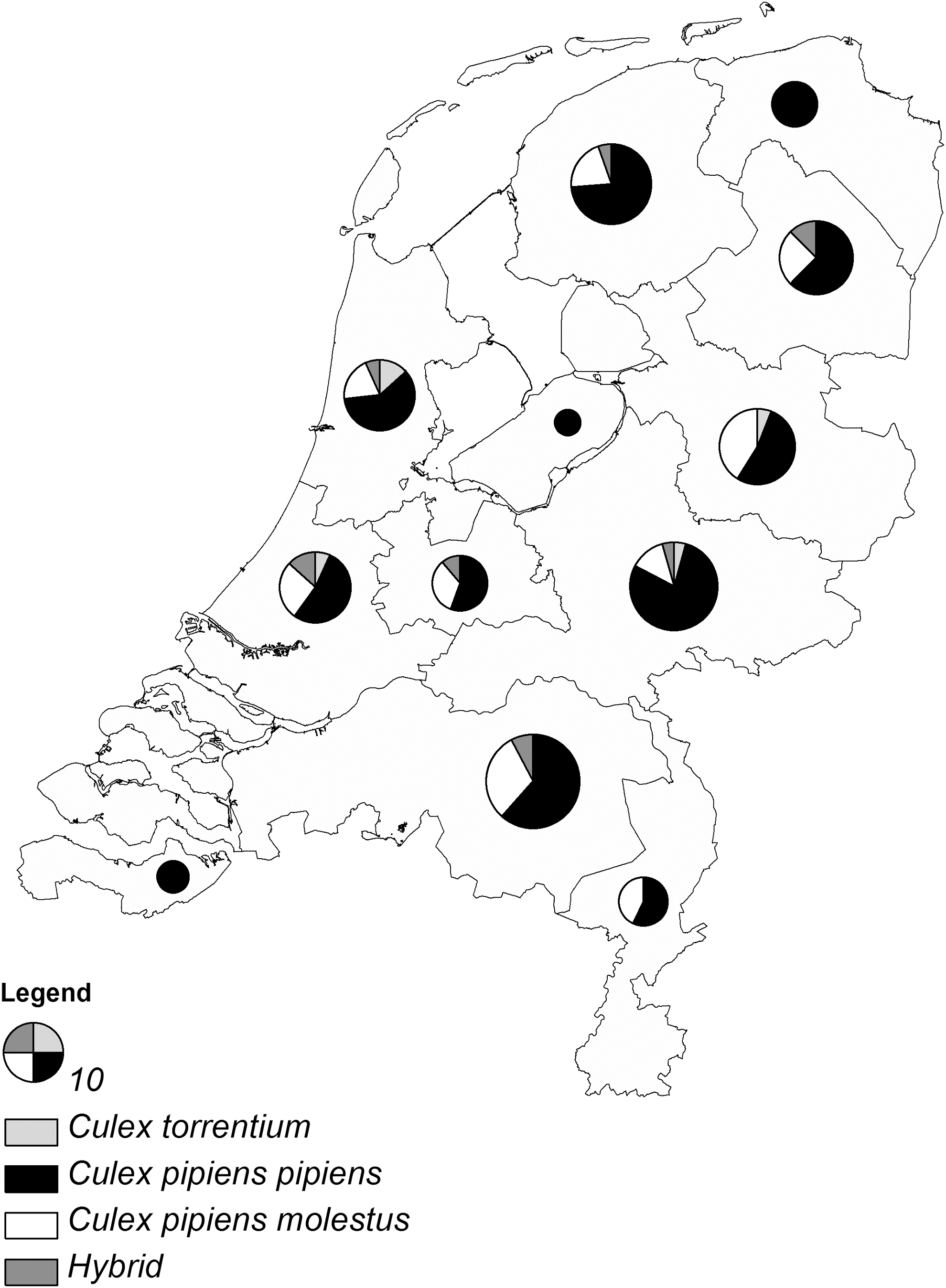
Culex mosquitoes collected outdoors with Mosquito Magnet Liberty Plus traps during the summers of 2011 and 2012 in The Netherlands. Relative composition of trapped Culex mosquitoes that were identified to the species and biotype level per province is shown. The size of each pie chart varies according to the number of analyzed samples.
Adult mosquito collection: Passive surveillance throughout The Netherlands
In total, 1563 mosquitoes were submitted during winter, from which 930 were identified as Culex mosquitoes. Out of 68 Cx. pipiens/torrentium mosquitoes that were blood-fed and had caused nuisance as reported by Dutch citizens from indoor locations during the winter of 2014, one was identified as pipiens (1.5%), 60 as molestus (88.2%), seven as hybrids (10.3%), and none as Cx. torrentium (0%) (Fig. 2A). In contrast, out of 82 nonnuisance mosquitoes that were submitted during winter, 58 were identified as pipiens (70.7%), 11 as molestus (13.4%), 12 as hybrids (14.6%), and one as Cx. torrentium (1.2%) (Fig. 2B). During summer, in total 581 mosquitoes were submitted, from which 397 were identified as Culex mosquitoes. From the total number of 89 mosquitoes that were blood-fed and had caused nuisance indoors during summer, six were identified as pipiens (6.7%), 62 as molestus (69.7%), 21 as hybrids (23.6%), and none as Cx. torrentium (0%) (Fig. 2C). In contrast, from the total number of 83 nonnuisance mosquitoes that were submitted during summer, 48 were identified as pipiens (57.8%), 30 as molestus (36.1%), five as hybrids (6.0%), and none as Cx. torrentium (0%) (Fig.2D).
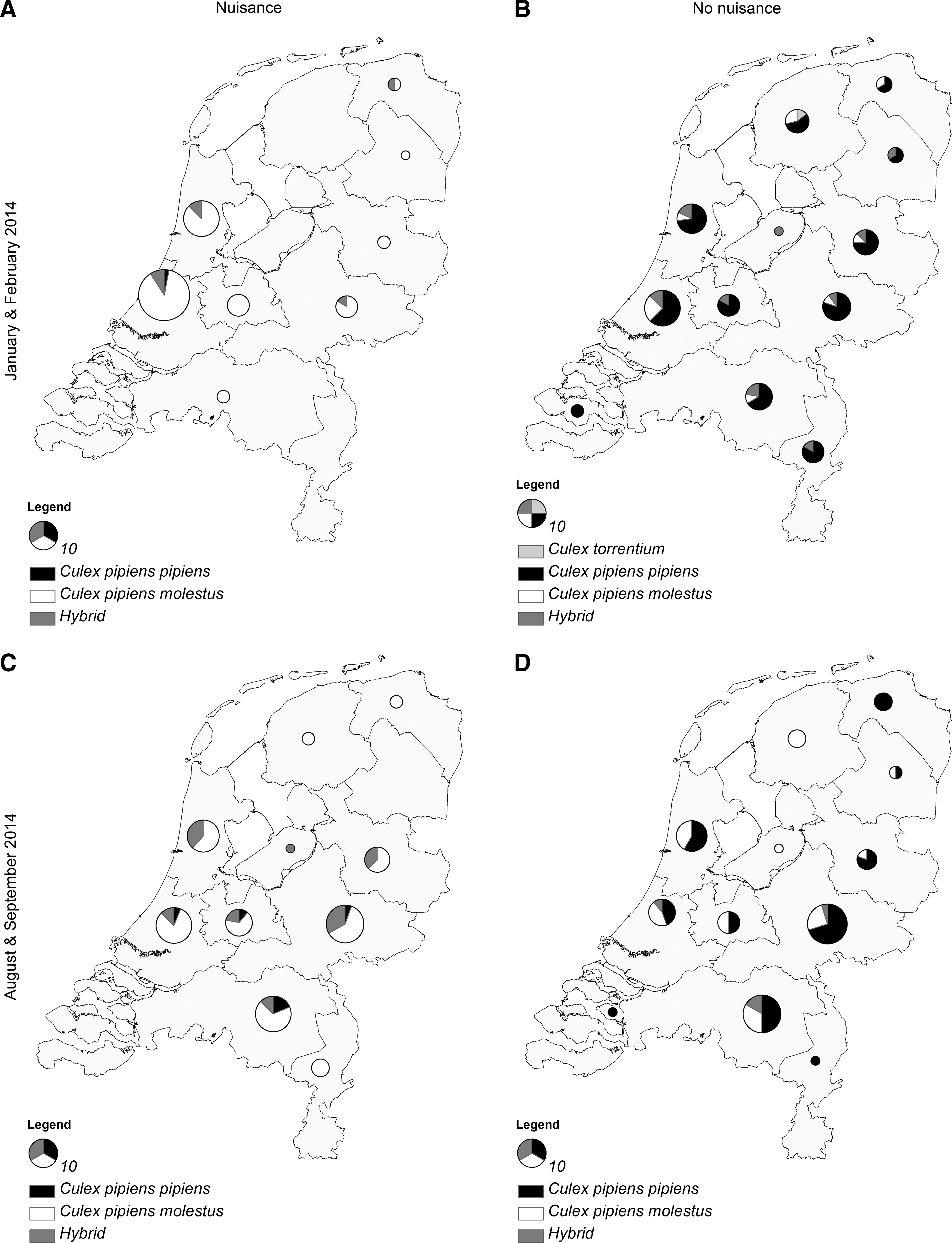
Culex mosquitoes collected indoors by Dutch citizens during the winter (January and February) and summer (August and September) of 2014 in The Netherlands. Relative composition of Culex mosquitoes per province, collected during January and February of 2014, that had caused nuisance and were engorged (
Larval and adult mosquito collection: In-depth study of the city of Amsterdam
From the 480 Cx. pipiens larvae that were analyzed, 436 were identified as pipiens (90.8%), 19 as molestus (4.0%), and 25 as hybrids (5.2%) (Fig. 3). These larvae were collected from a total number of 48 publicly accessible aboveground larval habitats, such as rainwater storm drains, man-made rainwater collections, and ditch water. In total, 26 larval habitats (54.2%) contained only biotype pipiens larvae, one larval habitat (2.1%) contained only biotype molestus larvae, one larval habitat (2.1%) contained only hybrid larvae, and 20 of the larval habitats (41.6%) contained a mixture of biotypes and/or hybrids.

Percentage of Cx. pipiens larvae and adults collected aboveground in Amsterdam. Cx. pipiens larvae (n = 480) collected at 48 larval habitats and Cx. pipiens adults (n = 161) collected at 20 locations with MM traps were identified to biotype level with real-time PCR. Dark-grey bars represent larvae, and light-grey bars represent adults. Values indicate exact percentages.
From the total number of 161 Cx. pipiens/torrentium adults that were collected in Amsterdam, 121 were identified as pipiens (75.2%), 26 as molestus (16.1%), and 14 as hybrids (8.7%) (Fig. 3). These results are similar to the nationwide results of active surveillance.
Discussion
The analysis of blood-fed specimens that had been submitted to our laboratory by members of the Dutch public suggests that both molestus and hybrids continue to feed indoors during winter and are mainly responsible for indoor bites year-round. Moreover, we found a high frequency of aboveground hybridization between the Cx. pipiens biotypes in The Netherlands. In contrast to the current paradigm that the molestus biotype is restricted to belowground habitats in northern Europe, we found that the pipiens and molestus biotypes occur sympatrically in aboveground locations.
In the view of biotypes occupying spatially separated habitats, it can be expected that hybridization is low (Chevillon et al. 1995, Fonseca et al. 2004). Hybrids of the two biotypes were described from The Netherlands in 2010, but they were exclusively reported from the belowground Amsterdam subway system (Reusken et al. 2010a). In the present nationwide study, we found that biotypes pipiens and molestus both occur aboveground (in mosquito traps, as well as inside houses of Dutch citizens), as is the case in southern European countries, with comparable hybridization rates ranging between 5% and 15% (Gomes et al. 2009, Amraoui et al. 2012, Gomes et al. 2013, Osório et al. 2014).
Not only adult molestus and hybrid mosquitoes were collected aboveground in this study, aquatic habitats containing molestus and hybrid larvae were also found to exist aboveground. Interestingly, relatively few larval habitats with biotype molestus were identified in comparison with biotype pipiens. The percentage of biotype molestus was higher for adults collected at the same locations in Amsterdam than for larvae. It is likely that a fraction of the typical underground biotype molestus habitats was not sampled, such as water collection basins in the subway (Reusken et al. 2010a). In addition to a bias in sampling of the larval populations, habitat characteristics and environmental conditions at trapping locations could have influenced the number and composition of collected adult mosquitoes (DiMenna et al. 2006, Giberson et al. 2007). An alternative explanation could be that biotype pipiens larvae are better competitors and, therefore, can outcompete biotype molestus larvae.
It remains unclear whether the molestus biotype has only recently expanded its niche from belowground to aboveground habitats or whether it has been aboveground for longer periods of time but was overlooked in previous studies. Locations where biotype molestus was found were distributed throughout the country. Therefore, we hypothesize that this is not a new development but that it has been aboveground for a longer period of time, or has always been present. This is supported by the collections of molestus from hotels and stagnant water in the United Kingdom by Willcox in the mid twentieth century (Willcox 1941). In the near future, we plan to evaluate population genetic structures in order to shed light on the status of the biotypes in northern Europe.
The lack of exponential increase in the amplification curves of well-characterized samples tested with the real-time PCR protocol developed by Rudolf et al. (2013) is probably due to competition between the “Culex pipiens all” and the “biotype pipiens” probes, as these were overlapping. Additionally, we found several single nucleotide mismatches between the biotype pipiens probe and sequences in GenBank. These issues were resolved by designing new primers and probes.
Passive surveillance with the help of citizens complements active surveillance tools that involve the use of traps (Kampen et al. 2015). Samples obtained through passive surveillance can provide specific insight into which mosquito species are present inside houses. Using this approach, our study revealed that biotype molestus was mostly responsible for biting nuisance, especially during winter. It is likely that during this season biotype pipiens has entered diapause and may thus be observed by citizens (Sim et al. 2008), but not experienced as nuisance. The numerous reports submitted via the website revealed that nuisance biting during winter is not uncommon. Such insights cannot be obtained by using traps. Traps, however, provide more insight into mosquito abundance, which is more difficult to achieve with passive surveillance.
To date, WNV has not been found to circulate among mosquitoes in a potentially high-risk area in The Netherlands, where both mosquitoes and wildlife are abundant (Reusken et al. 2010b). Because hybrids could play an important role in bridging WNV from birds to humans, the presence of year-round actively feeding hybrids in The Netherlands could constitute a risk if WNV were to be introduced in The Netherlands. Northwestern European Cx. pipiens mosquitoes have already been found to be competent vectors for WNV (Balenghien et al. 2007, 2008, Fros et al. 2015). The question, therefore, remains which factors, other than the presence of competent vectors, restrict WNV from establishing in northern Europe. A partial explanation may be that viremia levels in birds, such as European jackdaws, remain too low to be picked up by mosquitoes (Lim et al. 2014). Our future research efforts will focus on the impact of temperature on mosquito–virus interactions.
Conclusions
Biotype molestus and hybrids are active and feeding indoors during both summer and winter, and are mainly responsible for indoor biting nuisance. High levels of hybridization between Cx. pipiens biotype pipiens and molestus are observed in The Netherlands, which can be explained by sympatric occurrence of both biotype pipiens and molestus in aboveground habitats.
Footnotes
Acknowledgments
We thank the Dutch citizens who sent mosquito samples to our laboratory. Our thanks also go to Rens van den Biggelaar and Marian Dik for morphological identifications and colleagues from the Laboratory of Entomology, especially Linda Klein and Steffanie Teekema, for their help in processing all samples. We also thank Marcel Dicke and Alexandra Hiscox for providing comments on a previous version of this manuscript. This work is supported by the European 195 Community's Seventh Framework Programme (FP7 VECTORIE project number 261466) and the Uyttenboogaart-Eliasen Stichting (SUB.2014.05.15).
Author Disclosure Statement
No competing financial interests exist.