
Abstract
Chagas disease, which ranks among the world's most neglected diseases, is a chronic, systemic, parasitic infection caused by the protozoan Trypanosoma cruzi. Mepraia species are the wild vectors of this parasite in Chile. Host–parasite interactions can occur at several levels, such as co-speciation and ecological host fitting, among others. Thus, we are exploring the interactions between T. cruzi circulating in naturally infected Mepraia species in all areas endemic of Chile. We evaluated T. cruzi infection rates of 27 different haplotypes of the wild Mepraia species and identified their parasite genotypes using minicircle PCR amplification and hybridization tests with genotype-specific DNA probes. Infection rates were lower in northern Chile where Mepraia gajardoi circulates (10–35%); in central Chile, Mepraia spinolai is most abundant, and infection rates varied in space and time (0–55%). T. cruzi discrete typing units (DTUs) TcI, TcII, TcV, and Tc VI were detected. Mixed infections with two or more DTUs are frequently found in highly infected insects. T. cruzi DTUs have distinct, but not exclusive, ecological and epidemiological associations with their hosts. T. cruzi infection rates of M. spinolai were higher than in M. gajardoi, but the presence of mixed infection with more than one T. cruzi DTU was the same. The same T. cruzi DTUs (TcI, TcII, TcV, and TcVI) were found circulating in both vector species, even though TcI was not equally distributed. These results suggest that T. cruzi DTUs are not associated with any of the two genetically related vector species nor with the geographic area. The T. cruzi vectors interactions are discussed in terms of old and recent events. By exploring T. cruzi DTUs present in Mepraia haplotypes and species from northern to central Chile, we open the analysis on these invertebrate host–parasite interactions.
Introduction
C
To study the parasite–vector interactions of Mepraia at the individual, population, and species levels, we characterized parasites and vectors genetically. We previously examined the phylogeography of the insect vector on the basis of mitochondrial gene sequences (Campos et al. 2013b). The discrete typing units (DTUs) of Trypanosoma cruzi constitute reliable units for analysis for molecular epidemiology and experimental studies of evolution. The term “discrete typing unit” (Tibayrenc 1998) was proposed to describe sets of stocks that are genetically more similar to each other than to any other stock and are identifiable by common molecular markers sometimes referred to as “tags.” Experimental infections have demonstrated in pioneer studies that not all T. cruzi DTUs succeed in growing and completing their development in all triatomine species (Garcia et al. 1982, Azambuja et al. 1989, Mello et al. 1996, Urrea et al. 2005, Araújo et al. 2007, Mejía-Jaramillo et al. 2009, Vallejo et al. 2009, Araújo et al. 2014).
In this study, we characterized and gathered information about infection rates and the T. cruzi DTUs composition present in those naturally infected vectors and explored associations between these parameters. We performed genotyping by means of minicircle hybridization in T. cruzi infecting Mepraia species distributed along 2000 kilometerss of Chile. This method, which directly analyzes T. cruzi composition, thus avoiding bias due to selection by parasites culture, represents an effort to characterize the representative population of infective T. cruzi transmitted by the vector.
Materials and Methods
Insect collection and genetic characterization
M. gajardoi were collected during the austral spring–summer of 2008–2011 from coastal zones in northern Chile. Collection localities in Chile are shown in Figure 1. Information on the genetic characterization and haplotypes of individuals of each locality by means of cytochromes b and cytochrome oxidase subunit I is available (Campos et al. 2013b). Sites in the north have a coastal desert climate with less than 2 mm annual precipitation. These sites are extremely arid, have scarce plant cover, and include beaches with a mixture of rocks, pebbles, stones, and sand. Lizards, sea and carrion birds, and few wild rodents inhabit the collecting sites. Insects were collected from noon to 4:00 PM by two persons at each site.

Geographical location of insect collection places. Insects of the Mepraia genus were collected in different geographical areas, as indicated in northern and central Chile. Color images available online at
M. spinolai from central Chile were collected from areas of semiarid Mediterranean climate with most rainfall concentrated in winter (Di Castri et al. 1976). Several wild animals and very few birds inhabit the collection sites. They are rodents (Octodon degus, Phyllotis darwini, Abrothrix olivaceus), marsupials (Thylamis elegans), rabbits, caprines, and foxes. Captured bugs, most of them nymphs of different stages, were transported to the laboratory and kept separately in a climate chamber at 27°C with a relative humidity of 70% and a 14:10 hour light:dark photoperiod. Triatomines were analyzed immediately after collection by taking a fecal sample during or after feeding on noninfected Mus musculus. These field studies did not involve endangered or protected species. The bioethics committee on animal research in the Faculty of Medicine, University of Chile dated August 16, 2011, with the inscription “Protocol CBA # 0443 FMUCH” approved this study.
Amplification of T. cruzi minicircles
For parasitological analyses, we obtained fecal samples after feeding or rectal components from dead specimens. Fecal or intestinal contents were mixed with 100–200 μL of bidistilled water, centrifuged at 10,000 × g, and frozen at −20°C until the PCR was conducted. A 330-bp product indicated a positive result. The sample of triatomine fecal content was boiled for 15 min. Negative samples were evaluated for presence of inhibitors and DNA extraction was performed when they were present by means of the Favorgen kit (E.Z.N.A. Blood DNA Mini Kit) according to the manufacturer's instructions. Each PCR included positive and negative controls. A 1- to 5-μL sample of intestinal content was assayed in a mixture containing 121 oligonucleotides and 122 oligonucleotides, as described in Wincker et al. (1994).
T. cruzi genotyping of minicircle amplicons
For genotyping, PCR DNA blots were prepared by using 10 μL of each PCR product. Four T. cruzi clones (sp 104 cl 1, CBB cl 3, NR cl 3, and V195 cl 1), corresponding to Tc I, Tc II, Tc V, and Tc VI, respectively, were used to generate specific DTU probes. The construction of minicircle probes was performed as described (Veas et al. 1991). The PCR products were subjected to electrophoresis, transferred onto Hybond N+ nylon membranes (Amersham, Piscataway, NJ), and cross-linked by ultraviolet light for DNA fixation. After transferring the PCR products, four membranes were prehybridized for at least 2 h at 55°C. Each membrane was then hybridized with a DTU-specific probe labeled with 32P-α-d-adenosine triphosphate (32P-α-dATP) (1 × 106 cpm/membrane). After hybridization, membranes were washed under high-stringency conditions and then exposed in a Molecular Imager FX (Bio-Rad Laboratories, Hercules, CA). Variable amounts of amplicons (between 30 and 150 ng of DNA) were electrophoresed in 2% agarose gels prior DNA denaturing and later transferred to nylon membranes as described (Veas et al. 1991). Four copies of identical membranes containing DNA blots were hybridized against a panel of the DNA probes that recognize the specific DTUs Tcl (sp104cl1l), TcII (CBBcl3), TcV (NRcl3), and TcVI (V195cl1), which are the DTUs circulating in Chile as described (Barnabe et al. 2001, Rozas et al. 2007, Coronado et al. 2009, Arenas et al. 2012). The infection rates and the frequency of occurrence of each T. cruzi DTUs in M. gajardoi and M. spinolai were analyzed with a chi-squared test. The overall genotype distribution of T. cruzi DTUs between M. spinolai and M. gajardoi was analyzed with a G-test of goodness of fit.
Results
The specificity of the probes was evident in the hybridization assay performed with a control membrane containing the kinetoplast DNA (kDNA) of the four stocks used to generate them (Fig. 2). The T. cruzi infection rate was higher in M. spinolai compared with M. gajardoi (χ2 = 21.54, p < 0.001).

Probes control. Results of hybridization patterns of different T. cruzi stocks belonging to different DTUs. (
M. gajardoi were collected from extremely arid areas where mainly birds and lizards prevailed. M. spinolai circulates in semiarid areas where a diversity of vertebrate hosts exist rather than birds and lizards. The T. cruzi DTU composition circulating in Mepraia species vectors of Chile is diverse with 27 haplotypes (Campos et al. 2013b). However, they did not show a preferential spatial distribution, even though the geographic area studied is approximately 2000 km from north to south, except for a significant difference in TcI (χ2 = 71.38, p < 0.001, degrees of freedom [df] = 1) abundance between M. gajardoi and M. spinolai (Table 1).
(%) Percentage of insects infected in each locality.
Mepraia gajardoi distribution.
Mixed infections detected.
Mepraia spinolai distribution.
DTUs, discrete typing units of T. cruzi; NN, unidentified; ND, not determined.
The other T. cruzi DTUs (TcII, TcV, and TcVI) did not show a significant difference and were equally distributed over all the endemic regions studied and were equally present in vectors of the genus Mepraia. Infections were found to be single or mixed with two or more T. cruzi DTUs probes. Mixed infections are determined when the DNA blot hybridizes with more than one different probe (Fig. 3). Figure 3 also shows that unknown DTUs are circulating in some insects (samples 7 and 11). These cases are evidenced by the presence of a PCR band, without hybridization signals by any one of the four DNA probes tested; however, they do not represent more than 6.1% on average (Table 1).
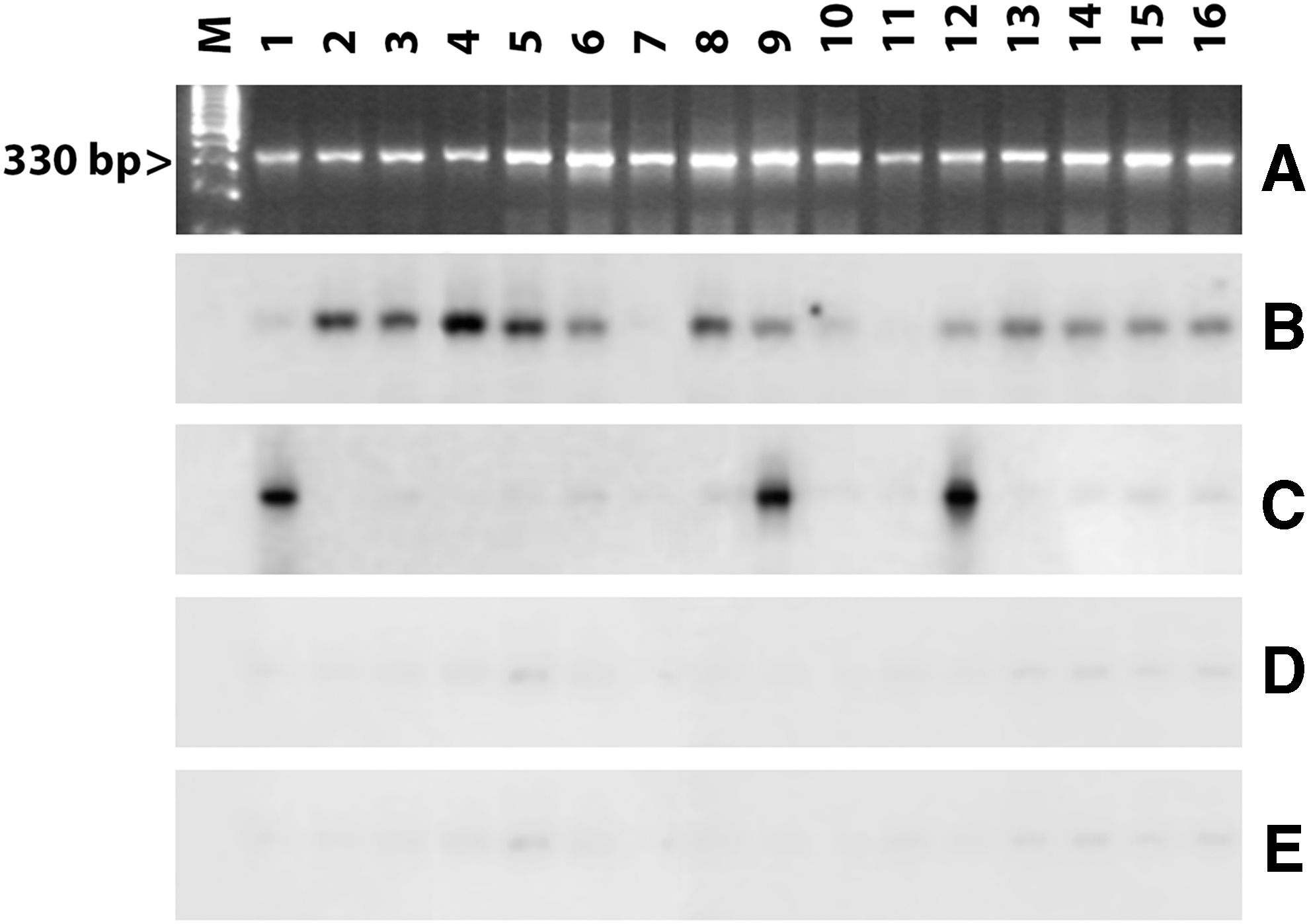
Representative results of T. cruzi DTUs by means of hybridization tests of Triatomine samples. (
The mixed infections are more prevalent when the infection rate is high, as found in M. spinolai. Infection rates detected in this species may be over 40%, and the composition of T. cruzi DTUs circulating in the different Mepraia species is summarized in Table 1. In a single infection, the sum of different DTUs and unidentified is the same as the number of insects infected; however, in the mixed infections, this number is higher, as shown in heavily infected localities such as Illapel and Colina. In some endemic areas such as the XV and I Regions in the extreme northern coastal ecoregion, the infection rate of M. gajardoi is low (10–35%); however, in others, such as Region IV and the Metropolitan Region, the infection of M. spinolai can reach up to 54%, allowing detection of mixed infection with different combinations of T. cruzi DTUs.
The most frequent T. cruzi DTUs combinations were TcI with TcII, TcI with TcV, TcI with TcVI, and TcI with TcII and TcVI (not shown). Table 1 shows that M. gajardoi carries mainly single infections with TcI and TcII as the most abundant DTUs in Playa Corazones, Caleta Vitor, and Caleta Camarones; however, M. gajardoi from two localities (Rio Seco and San Marcos) separated by ∼200 km at the South in the North ecoregion had single infections of DTUs TcV and TcVI as the most frequent. Mepraia spp. from localities 1000 km farther south correspond to another species named M. parapatrica. The T. cruzi DTUs from this area were only studied in one locality (Caleta Zenteno). The most abundant T. cruzi DTU was TcI. Finally, in the last species corresponding to M. spinolai from several localities in the center of Chile, some of the triatomines were found to be heavily infected (up to 50%). The most frequent T. cruzi DTUs in this area, which includes the coast and interior valleys up to 2000 meters, are TcI, TcII, TcV, and TcVI. The overall genotype distribution of T. cruzi DTUs was significantly different between M. spinolai and M. gajardoi (G = 48.52, p < 0.001, df = 3).
Discussion
Mepraia species show a heterogeneous population genetic structure across the latitudinal span of Chile. M. gajardoi and M. parapatrica inhabit the hyperarid coastal areas of northern Chile, whereas M. spinolai inhabits coastal and internal central areas of Chile up to 2500 meters altitude. Studies comparing rates of nucleotide substitution of mitochondrial genes and times of most recent common ancestor suggest that M. spinolai diverged about 2.9 mya (Campos et al. 2013b). This contrasts with the temporal scale of divergence for the most ancient extant T. cruzi DTUs TcI and TcII, about <1–4 mya (Flores-Lopez et al. 2011, Lewis et al. 2011). M. gajardoi and M. parapatric both have diverged more recently, 1.0 mya and 0.7 mya, respectively (Campos et al. 2013b). Higher infection rates and presence of mixed infections were found in M. spinolai rather than in M. gajardoi. Clearly M. spinolai feeds on a variety of hosts (mammals prevail over lizard and birds). Meanwhile, M. gajardoi feeds more frequently on the last two. One explanation for the finding of lower infection rates in M. gajardoi could be that some of the blood donor vertebrates such as birds are refractory to T. cruzi infections and interrupt T. cruzi transmission (Kierszenbaum et al. 1976). Interestingly, insects of Caleta Vitor analyzed after two consecutive feedings with M. musculus resulted in infection rates over 65% (Egaña et al. 2014).
Meanwhile, T. cruzi DTUs circulating in the sylvatic and domestic cycles of the central area of Chile are TcI, TcII, TcV, and TcVI (Rozas et al. 2007, Coronado et al. 2009). Probably some unknown subgroups of TcI and TcII (different than sp104cl 1 and CBBc13) are also present, because these DTUs are the oldest and most diverse (Arenas et al. 2012). However, the unknown genotypes detected here do not represent a large number, because these were not found at a rate of more than 6.1% on average. T. cruzi DTUs TcIII and TcIV have not been found in Chile using isoenzyme analyses in a large number of T. cruzi strains isolated from sylvatic and domestic hosts (Miles et al. 1984, Barnabe et al. 2001). However, the most frequently found T. cruzi DTUs in Mepraia species are the ancient TcI and TcII, and the least frequent are the hybrids TcV and TcVI, which diverged about 0.5–0.7 mya (Flores-Lopez et al. 2011) or even less, e.g., 0.06 mya (Lewis et al. 2011).
Even though the hybrid T. cruzi DTUs are more recent than the Mepraia species, it seems they are well adapted in these invertebrates. They were found in M. gajardoi of Rio Seco and San Marcos, where TcVI was the most represented DTU. This observation suggests that specific T. cruzi DTUs in some geographic areas are not due to strict associations with the invertebrate host, but rather to the opportunity to infect with specific T. cruzi DTUs. The presence of TcVI in these two wild localities is related to the proximity with domestic animals mainly dogs, fisherman, and algae collectors. Previous results of T. cruzi DTUs circulating in Mepraia species resulted in infection with TcI, TcII, TcV, and TcVI DTUs (Coronado et al. 2009, Toledo et al. 2013). However, they present temporal variations, because in Illapel the frequency of these T. cruzi DTUs varied in favor of TcI as the most represented, with very few TcII, TcV, and TcVI, as previously reported (Coronado et al. 2009). Two years later in the same locality, a high infection rate and similar presence of the four DTUs were detected, as reported here. More recently in Illapel, the most representative T. cruzi DTUs mixtures were TcI and TcV; the mixture of TcI with TcV and single infections with TcVI were not found (Botto-Mahan et al. 2015).
The other locality with a high presence of mixed infection is Colina, where the combinations of TcI with TcII, TcII with TcVI, and TcI with TcII and TcVI were the most represented. We suggest that the frequent association of mixed infections in M. spinolai can be explained by the high diversity of mammals carrying different T. cruzi DTUs. The overall genotype distribution of T. cruzi DTUs between M. spinolai and M. gajardoi was significantly different. This can be explained by the high frequency of TcI in M. spinolai, which skews the overall distribution analysis.
T. cruzi transmission to humans in Chile seems to be old, as suggested by evidence of Chagas disease in mummies of the far north coastal Chinchorro culture (Aufderheide et al. 2004). These estimates are of 9000 years, when probably only the indigenous M. gajardoi existed in that area. Recent studies of tissue of mummies of Chinchorro and the 2400- to 3000-year-old Alto Ramirez culture determined that Tc I, Tc II, and Tc IV DTUs were present at those times (Guhl et al. 2014). The finding of TcIV in ancient humans opens the question of whether this DTU is currently present in Mepraia spp. as an unknown DTU. All of these unknown DTUs can be variants of TcI and TcII, or TcIV, that is, they are ancient T. cruzi DTUs. The other triatomine present in Chile so far controlled is T. infestans, which is thought to have dispersed to southern endemic areas from an initial sylvatic focus in Bolivia or Argentina (Cortez et al. 2010, Torres-Perez et al. 2011). This event probably was recent, not older than the initiation and expansion of the Inca Empire in the Southern Cone (Solari 2011), or 2000 years ago according to dating of mummies infected with T. cruzi in San Pedro de Atacama, Chile, which is one highland area far away from the coast and close to the Bolivian border (Ferreira et al. 2000).
It is thought that, because triatomine species contain innate defenses against some T. cruzi strains, such as hemolytic factors and also other microorganisms in the digestive tract, these hemolysins and microbiota may discriminate between different T. cruzi DTUs (García et al. 2010). There is a tendency of local insect vectors to be more susceptible to strains of T. cruzi from the same geographical areas, as demonstrated with xenodiagnosis studies in humans (Perlowagora et al. 1988). Even though some exceptions exist (Araújo et al. 2008), experiments to demonstrate the differential susceptibility of M. spinolai and T. infestans to detect T. cruzi from naturally infected O. degus of Chile determined that M. spinolai has a higher parasite amplification capacity than T. infestans (Campos et al. 2007).
In nature, the circulating T. cruzi parasites are mixed populations, as observed here using a direct method to genotype T. cruzi, especially in highly infected vectors. The evidence to demonstrate co-speciation between parasite and hosts relies on phylogenetic analyses of host and parasite genome sequences, an obligatory methodology that can favor parasite artificial selection during T. cruzi culture, as determined by Deane et al. (1984), or DNA sequencing from a polyclonal infection. Therefore, in these circumstances, it is very difficult to demonstrate host–parasite co-speciation. Efforts to find associations between two Mepraia species and a T. cruzi DTU in a longitudinal area of 2000 km, as shown in this study, seem unsuccessful. The parasite–host association is not restrictive, because different T. cruzi DTUs can co-exist in each host. The epidemiological impact of our results is that two Mepraia species from any latitude may carry the same four most represented DTUs (TcI, TcII, TcV, and TcVI). Although T. cruzi and Mepraia species do not show co-speciation, it is clear that Chagas disease is dependent on a high degree of interactions between triatomines and parasites. However, these interactions can be recent, as seems to occur as shown here with M. gajardoi and T. cruzi DTUs TcVI, which diverged very recently. Finally, since not all triatomine species develop different T. cruzi DTUs, this suggests that triatomines of a different genus rather than species of the same genus are differently adapted to each T. cruzi DTU (Mejía-Jaramillo et al. 2009, Araújo et al. 2014). The different microbiota present in triatomine guts of different genera seem to contribute to this interesting and epidemiologically relevant parasite–invertebrate interaction (Da Mota et al. 2012).
Conclusions
T. cruzi infection rates of M. spinolai were higher than in M. gajardoi, with the same presence of mixed infection with more than one T. cruzi DTU. The same T. cruzi DTUs (TcI, TcII, TcV, and TcVI) were found circulating in both vector species, even though TcI resulted in not being equally distributed. These results suggest that T. cruzi DTUs are not associated with any of the two genetically related vector species nor with the geographic area. The T. cruzi vectors interactions were discussed in terms of old and recent events.
Footnotes
Acknowledgments
This work was supported by FONDECYT-Chile 1120122 to A. Solari and 3150289 to R. Campos-Soto.
Author Disclosure Statement
No competing financial interests exist.