
Abstract
In previous studies, we have demonstrated that inoculation with a Trypanosoma cruzi marinkellei (avirulent RM1 strain) was able to reduce parasitemia in mice challenged with T. cruzi, although it was not able to prevent histopathological lesions. Th1 response stimulation by immunization is necessary for T. cruzi infection control, but the resistance is also dependent on immunoregulatory mechanisms, which can be induced by adjuvants. Thus, we evaluated whether inoculation of T. cruzi marinkellei associated with administration of different adjuvants would be capable of inducing different patterns of immune response to maximize the immune response against T. cruzi (virulent Romildo strain) infection. Two hundred eighty nonisogenic mice were divided into 14 groups according to the immunization scheme and the subsequent challenge with virulent Romildo T. cruzi strain. Nonimmunized groups and animals inoculated without adjuvants were also included. Immune protection was not observed with Th2 adjuvants (incomplete Freund's adjuvant [IFA] and Alum) due to high parasitemia. Th1/Th2-polarizing adjuvants also did not induce immune protection because inulin was unable to maintain survival, and immune-stimulating complexes induced intense inflammatory processes. Animals sensitized with RM1 strain without adjuvants were able to reduce parasitemia, increase survival, and protect against severe histological lesions, followed by adequate cytokine stimulation. Finally, our results demonstrate that the early and balanced IFN-γ production becomes critical to promote protection and that Th1 adjuvant elicited a controversial infection control due to increased histopathological damage. Therefore, the host's immunomodulation remains one of the most important challenges in the research for effective protection against T. cruzi infection. Similarly, the identification of protective antigens in the RM1 strain of T. cruzi marinkellei may contribute to further studies on vaccine development against human Chagas disease.
Introduction
M
The high prevalence of Chagas disease in Latin America and its current expansion to developed countries (Hotez et al. 2013), side effects of actual therapies, lengthy treatments, low efficiency, and high costs of clinical treatment and assistance at the chronic stage have motivated studies on immune interventions (Urbina and Docampo 2003, Dumonteil et al. 2012). Other factors that may also justify these studies are the resistance of vectors to insecticides currently used, the risk of other modes of nonvectorial transmission (oral, congenital, blood transfusion, and laboratory accident), and the instability of control programs (Camargo 2009, Bonney 2014). The choice of immune interventions is critical and has been investigated by many researchers around the world since it is already known that the immune response to T. cruzi has a very positive effect on the healing process or reduction of injury resulting from infection. In other words, the immune response against T cruzi has been central to understanding the disease and hence its cure.
Since the 1960s, research has aimed to investigate the immune protection against infection using mechanisms of cross protection with attenuated or inactivated strains of trypanosomes (Menezes 1969, McHardy 1977), live genetically modified parasites (Perez Brandan and Basombrio 2012, Sanchez-Valdez et al. 2014), recombinant proteins or plasmids containing genes of T. cruzi (Bhatia and Garg 2008, Rodriguez-Morales et al. 2013), and DNA sequences inserted in viral vectors (Duan et al. 2009, Barbosa et al. 2013). Several antigen candidates have been chosen for further study, such as cruzipain, trans-sialidase, the protein regulating the complement, the amastigote surface protein, and a paraflagellar structural protein (Garg and Tarleton 2002, Vasconcelos et al. 2004, Dumonteil 2007). The search for effective immunogens has extended to cDNA libraries constructed from T. cruzi in specific stages, aiming at the identification of new antigens or even the composition of a multigenic vaccine in silico (Tekiel et al. 2009).
The complexity of the parasite–host relationship during infection by T. cruzi may characterize Chagas disease as a consequence of the host's failure to eliminate the infection, resulting in tissue damage induced by the parasite and mediated by the immune system. Thus, identifying protective antigens that do not promote an exacerbated inflammatory response against the parasite is a major step for obtaining an effective vaccine. In this sense, adjuvants also play an essential role driving CD4+ T-cell differentiation into a specific Th phenotype (Brummelman et al. 2015), influencing the type of response unleashed against the parasite.
In the early stages of contact between T. cruzi and the host, macrophages and dendritic cells lead to the activation of the immune system, which is evidenced by increased serum levels of chemokines and proinflammatory cytokines (Gazzinelli et al. 2006, Tarleton 2007). Early production of IL-12, IFN-γ, and TNF-α becomes a critical event in directing T cells to the Th1 phenotype leading to resistance to T. cruzi infection (Dutra et al. 2005, Machado et al. 2012a).
Immunization studies involving strains of trypanosomatids other than T. cruzi, such as Trypanosoma rangeli (Zuniga et al. 1997, Palau et al. 2003, Basso et al. 2014), Leptomonas pessoai (Souza et al. 1974), and Phytomonas serpens (Bregano et al. 2003, Pinge-Filho et al. 2005), are relevant not only from the antigenic point of view but also for clarifying the mechanisms of the host's immune response activation.
In a previous study by our group, we observed that inoculating mice with the RM1 strain of T. cruzi marinkellei, isolated from the bat Phyllostomus hastatus (Bento; Ramírez, unpublished data), resulted in an intense reduction of parasitemia after the challenge with two virulent strains of T. cruzi. However, despite this partial protection, we observed severe histopathological lesions with a strong presence of eosinophils in the tissue (Nascentes et al. 2008, 2010).
Therefore, due to difficulties in obtaining complete protection against T. cruzi infection and possible autoimmune phenomena triggered by the parasite and/or its products, it is necessary to clarify how inoculation with nonpathogenic strains of trypanosomatids interferes with the immune response of the host and if an appropriate adjuvant may result in correct modulation of immune response during a challenge with virulent strains of T. cruzi. Thus, the aim of this study was to observe whether immunization with a strain of Trypanosoma cruzi marinkellei isolated from bats, combined with adjuvants of different immune profiles, is able to protect nonisogenic mice against infection induced by a pathogenic strain of T. cruzi.
Materials and Methods
Animals
Two hundred eighty nonisogenic male mice (Swiss lineage, 4 to 8 weeks old) with weights ranging from 30 to 40 g were divided into 14 groups according to the immunization strategy and T. cruzi challenge. The animals were kept under suitable temperature and humidity conditions, and water and food were available ad libitum. The present work was approved by the Animal Use Ethics Committee of the Federal University of Triângulo Mineiro (Protocol 71) according to the Ethical Principles in Animal Research adopted by the National Council for Animal Experiments Control (CONCEA).
Trypanosomatid strains and adjuvants
The T. cruzi marinkellei RM1 strain, which was isolated from a Phyllostomus hastatus bat and is not, in nature, able to infect mice (Nascentes et al. 2008, 2010), was used to sensitize the animals. The virulent Romildo strain of the T. cruzi II phylogenetic group used to infect the animals was isolated at the Parasitology laboratory of Federal University of Triângulo Mineiro from a cardiodigestive form of Chagas disease (Machado et al. 2012b). Adjuvants used in association with the RM1 strain were chosen according to their immunogenic capacity at eliciting a cellular immune response (complete Freund's adjuvant [CFA]; SIGMA), a humoral immune response (incomplete Freund's adjuvant [IFA], and aluminum hydroxide gel, Alum; SIGMA), or both Th1/Th2 phenotypes (inulin, SIGMA; and immune-stimulating complexes [ISCOMs]). ISCOMs were prepared according to Pham et al. (2006), using cholesterol, phosphatidylcholine, and purified saponin (Pham et al. 2006).
Experimental design
The animals were divided into 14 groups of 20 animals each according to the immunization by adjuvant (CFA, IFA, Alum, Inulin, and ISCOMs) in combination with the RM1 strain of T. cruzi marinkellei and by the subsequent challenge with the Romildo strain of T. cruzi. The groups included mice immunized without adjuvant and nonimmunized groups. Seven groups were challenged, and seven were kept as noninfected controls. The animals were sensitized by subcutaneous injection with 100 μL containing 1.0 × 106 trypomastigotes of the RM1 strain emulsified with saline or adjuvant. Twenty-one days after immunization, mice were challenged with 5.0 × 103 trypomastigotes of the Romildo strain of T. cruzi through intraperitoneal injection (Fig. 1).
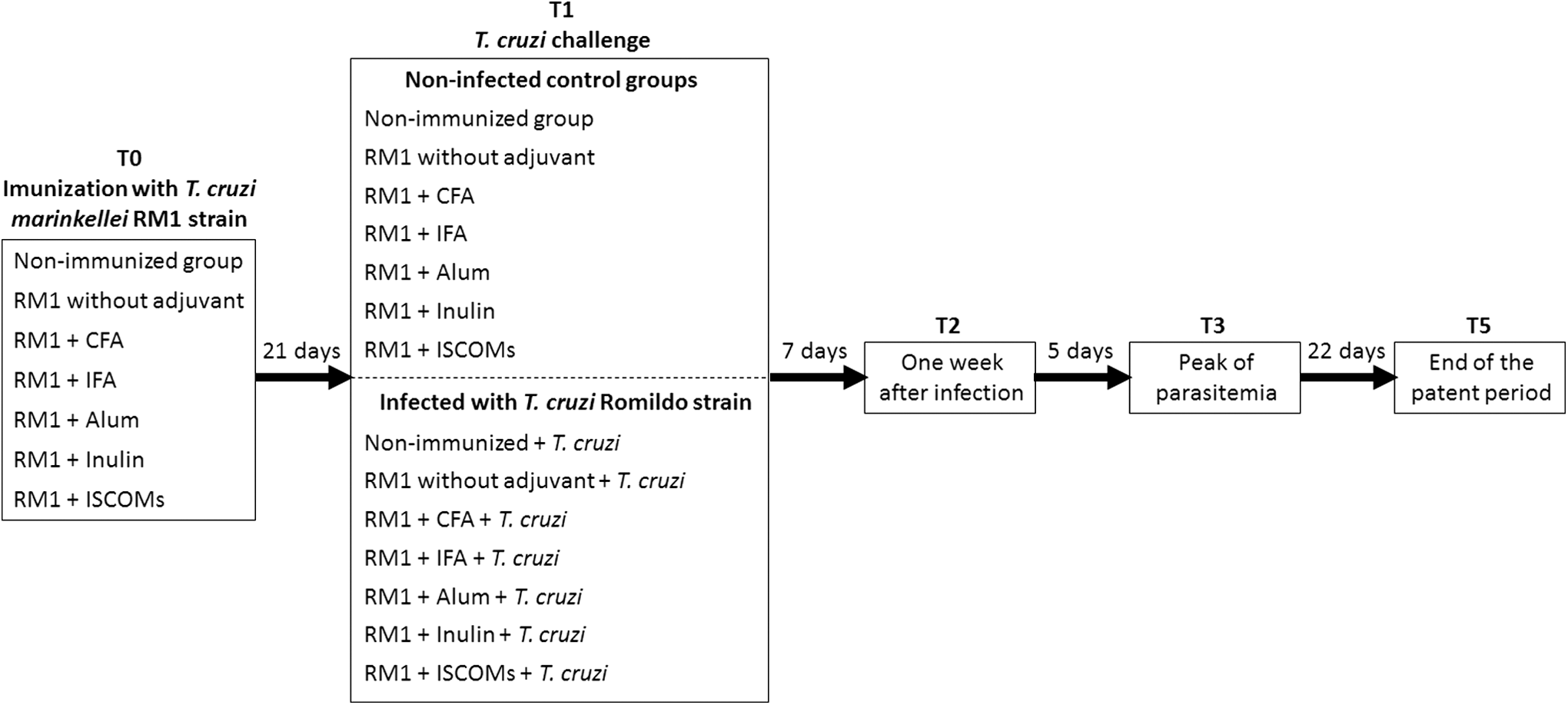
Flowchart of the experimental design. The animals were divided into 14 groups according to the immunization strategy and subsequent challenge with the Romildo strain of Trypanosoma cruzi. Blood samples were collected at the periods, T0 (before immunization), T1 (before infection), T2 (1 week after infection), T3 (at the peak of parasitemia), and T5 (at the end of the patent period).
Parasitemia, histopathology studies, and survival
Parasitemia was measured daily in each group of 10 animals throughout the patent period (Brener 1962). At the end of the patent period, which was 35 days postinfection, 10 animals of each group were euthanized by cervical dislocation under ether anesthesia. Tissue specimens from the heart, skeletal muscles of the quadriceps, the bladder, and gastrointestinal joints (gastroesophageal, gastroduodenal, and ileocecal) were collected and two 3–4-μm sections of each organ per animal were processed with hematoxylin and eosin staining. We investigated amastigote nests in the tissue, and the inflammatory processes were classified with a semiquantitative gradual scale (between 0 and 7: −, −/+, +/−, +, +/++, ++, ++/+++, +++), considering the intensity of inflammation (absent, mild, moderate, and severe) and its extension (focal or diffuse), as previously described (Ben Younes-Chennoufi et al. 1988, Garcia et al. 2006, Santos et al. 2007, Frank et al. 2008). The mortality of the animals was recorded during a period of 120 days of infection, which corresponds to the acute phase of infection.
Quantification of cytokine levels
A CBA Mouse Th1/Th2 Profile Kit, including IL-2, IL-4, IL-5, IL-10, IFN-γ, and TNF, and CBA Mouse IL-10, IL-12p70, and IL-17A Flex Sets (BD Biosciences) were used to measure cytokine levels in plasma, following the manufacturer's instructions. Quantification was performed before immunization (T0), before infection (T1), 1 week after infection (T2), at the peak of parasitemia (T3), and at the end of the patent period (T5). Data were acquired in a Becton Dickinson FACSCanto flow cytometer and analyses were performed using FCAP ArrayTM v.1.0.1 for Windows.
Statistical analysis
Comparisons among the different groups were carried out using analysis of variance (ANOVA), followed by the Bonferroni's post hoc test, for parametric data or Kruskal–Wallis, followed by Dunn's test, for nonparametric data. The evolution of cytokine levels between intervals of blood collection was assessed using ANOVA for repeated measurements or Friedman test, according to parametric or nonparametric distribution, respectively. The cumulative survival was analyzed by the Kaplan–Meier curve and the groups were compared through the log-rank test and calculation of hazard ratio. The statistical analysis was accomplished using Statistica 8.0 (Statsoft, Inc., 2008) and GraphPad Prism 5.0 (GraphPad Software, Inc., 2007), and p-values <0.05 were taken to be significant.
Results
The presence of T. cruzi was detected by analysis of parasitemia, which allowed observing the behavior of Romildo T. cruzi strain in immunized groups with different adjuvants associated with T. cruzi marinkellei RM1 strain and in the nonimmunized group. The curves of parasitemia revealed that the patent period was similar among the infected groups (p = 0.062), ranging between 21 and 34 days. It is noteworthy that animals immunized with RM1 strain or those immunized with RM1 strain associated with CFA, Inulin, and ISCOMs had lower peaks of parasitemia than did both the animals immunized with standard Th2 adjuvants (IFA and Alum) and nonimmunized animals (p < 0.001) (Fig. 2A, B).
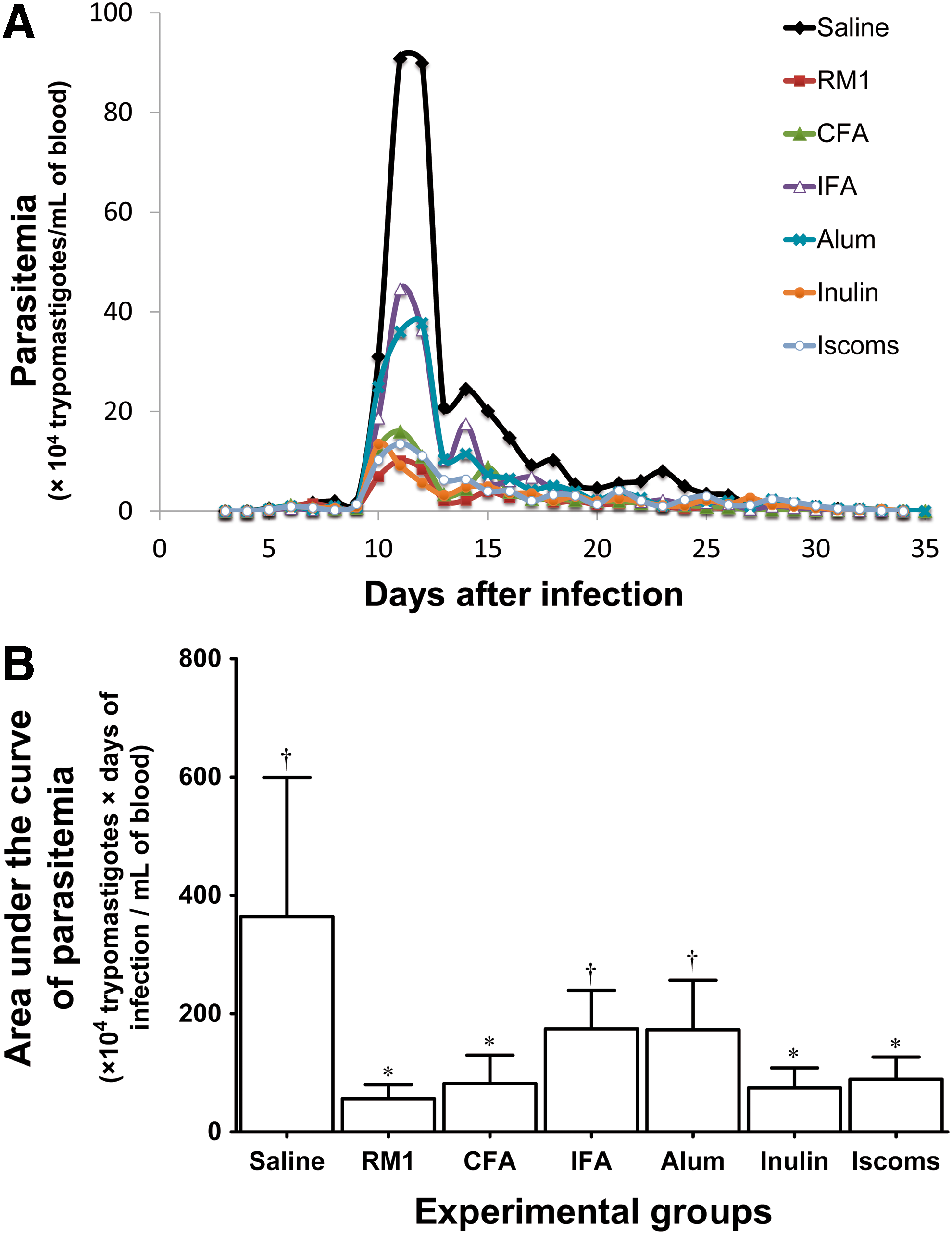
The behavior of T. cruzi Romildo strain parasitemia in animals immunized with different adjuvants.
The evaluation of animal survival in all groups was accomplished for 120 days after infection and showed that the Romildo strain of T. cruzi did not induce high mortality in mice during the acute phase of infection because survival ranged between 65.0% and 100.0%. However, comparing different immunization strategies, it was observed that animals sensitized with standard Th2 (IFA and Alum) adjuvants or with Inulin (Th1/Th2) had a lower survival rate compared with other animals (p = 0.002) (Fig. 3). In addition to the Kaplan–Meier curve, which shows the cumulative survival according to observation time, survival analysis allows for calculating the rate of death among the different groups (Hazard ratio). We attempted to analyze the impact of the adjuvant in animal mortality using animals that were sensitized only with RM1 strain as reference. Mice immunized with IFA, Alum, or Inulin had significantly lower survival rates corresponding to 10.06 (2.24–45.21), 8.42 (1.16–61.02), and 11.57 (1.93–69.39) respectively, than animals immunized with the RM1 strain in the absence of adjuvant (Fig. 3).
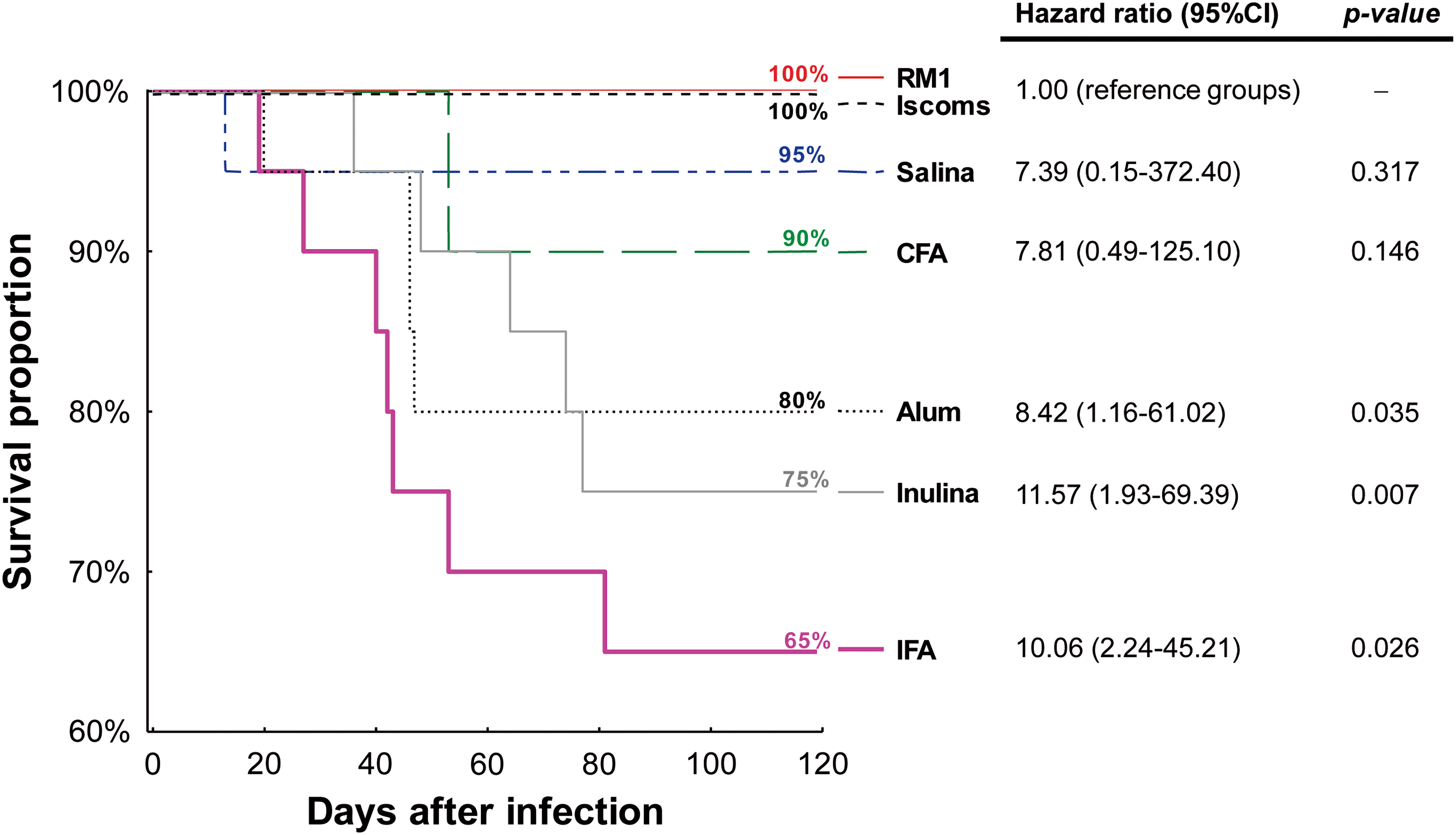
Kaplan–Meier plots of survival after 120 days of infection in animals immunized with different adjuvants and challenged with T. cruzi Romildo strain. p-Values for log-rank analyses and hazard ratios (95% confidence interval) for mouse survival were estimated taking the animals immunized with RM1 strain without adjuvants or those immunized with RM1 strain associated with ISCOMs (100% survival). Color images available online at
Regarding the histological finding, we investigated the presence of amastigote nests and inflammatory infiltrates in the heart tissue, skeletal muscles of the quadriceps, bladder, and gastrointestinal joints (gastroesophageal, gastroduodenal, and ileocecal). Comparing parasitism among different immunization strategies, there was no significant difference between groups in the percentage of parasitized mice. Despite the lack of statistical significance, it should be noted that animals immunized with Alum presented amastigote nests in almost all of the assessed tissues. With regard to inflammatory infiltrates, the analysis of quadriceps skeletal muscle presented the unique and significant differences between groups according to the immunization strategy. It is important to further emphasize that the group immunized with the RM1 strain with no adjuvant presented less skeletal muscle inflammation than did the groups that received saline, Alum, or ISCOMs (Fig. 4).
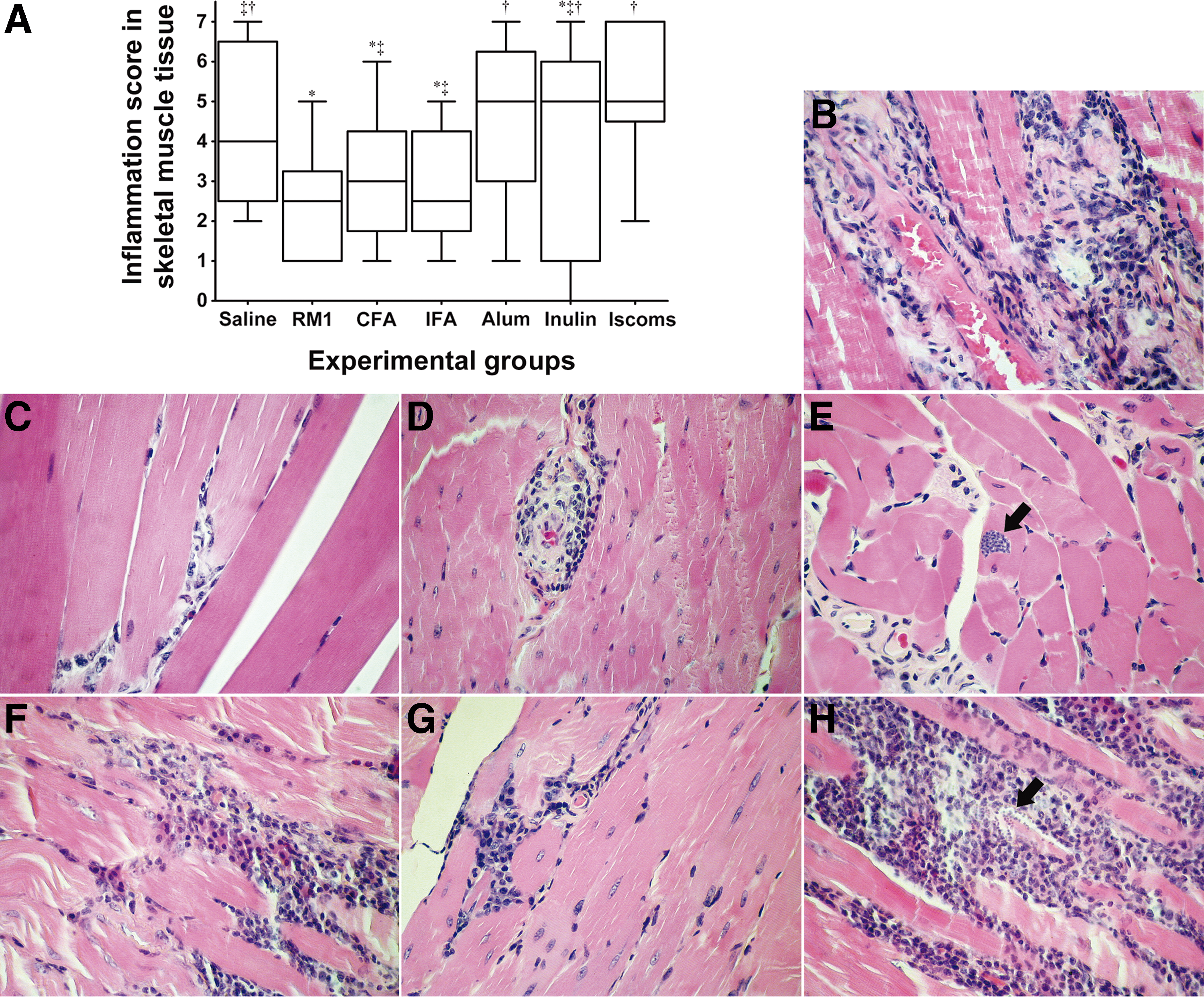
Inflammatory process in skeletal muscle of the quadriceps of animals immunized with different adjuvants and infected with T. cruzi Romildo strain.
Aiming the characterization of immune response induced by immunization and its comparison with nonimmunized animals, the cytokine profiles produced before and after infection by Romildo strain of T. cruzi (T0 to T5) were evaluated. Our results showed that serum levels of IL-2, IL-4, and IL-17 were lower than 10.0 pg/mL, with no significant differences observed between the studied groups (data not shown). Most mice showed no detectable levels of IL-10 or IL-12 (data not shown).
The animals challenged with the Romildo strain of T. cruzi, regardless of the immunization method, and the nonsensitized animals showed higher serum levels of TNF-α than did control animals both at times T3 and T5 (Fig. 5a). Furthermore, there was no difference between groups immunized with different adjuvants and nonimmunized animals.
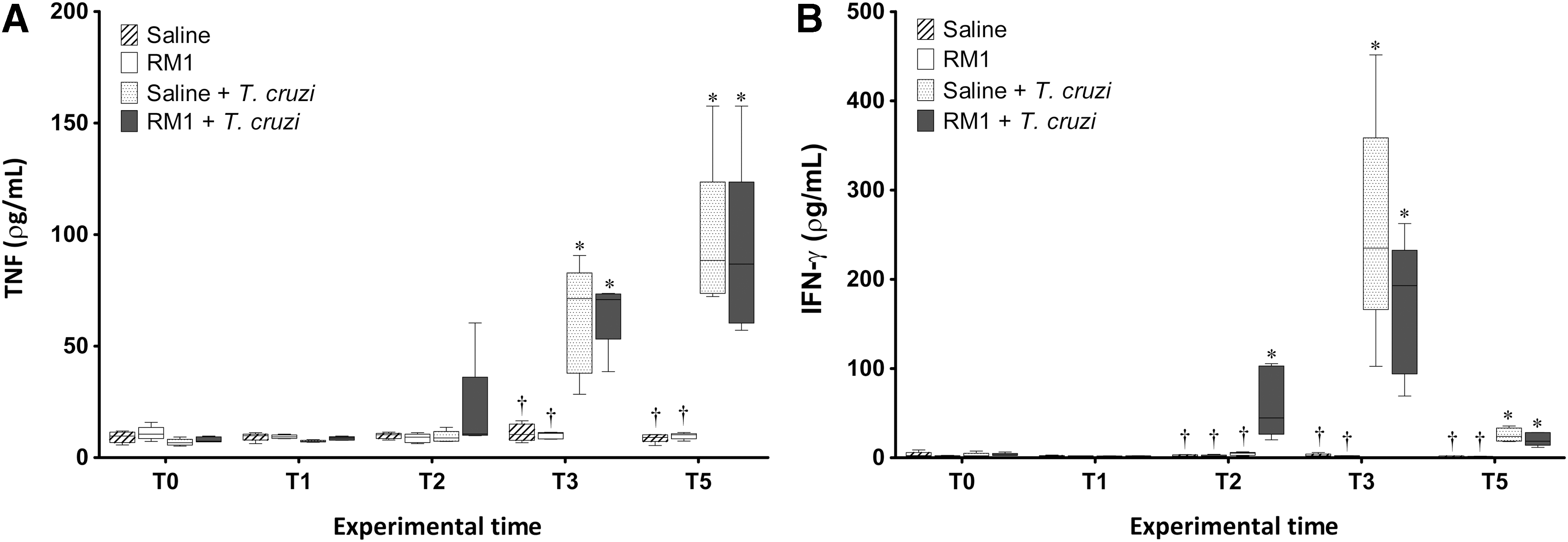
TNF
Infection by the Romildo strain of T. cruzi also led to an increase in the serum levels of IFN-γ in all animals, regardless of immunization, and the concentration of this cytokine peaked concurrently with the peak of parasitemia (T3). However, the levels observed in animals that were previously immunized with the RM1 strain associated with adjuvants were higher than in the nonimmunized infected group at time T2, 1 week after infection (Fig. 5b).
Animals sensitized with standard Th2 adjuvants (IFA and Alum) showed higher levels of IL-5 1 week after infection (T2) that were further elevated at the parasitemia peak (T3) (Fig. 6). At T2, nonimmunized animals and those immunized with IFA and Alum adjuvants had lower levels of IFN-γ than did animals immunized with CFA, Inulin, ISCOMs, and with RM1 strain without adjuvant. In contrast, at T3, animals immunized with IFA and Alum had higher IFN-γ cytokine concentrations in relation to the other animals, particularly compared with the groups sensitized with Inulin, ISCOMs, and with RM1 strain without adjuvant. However, these animals displayed delayed production of IFN-γ (Fig. 7).
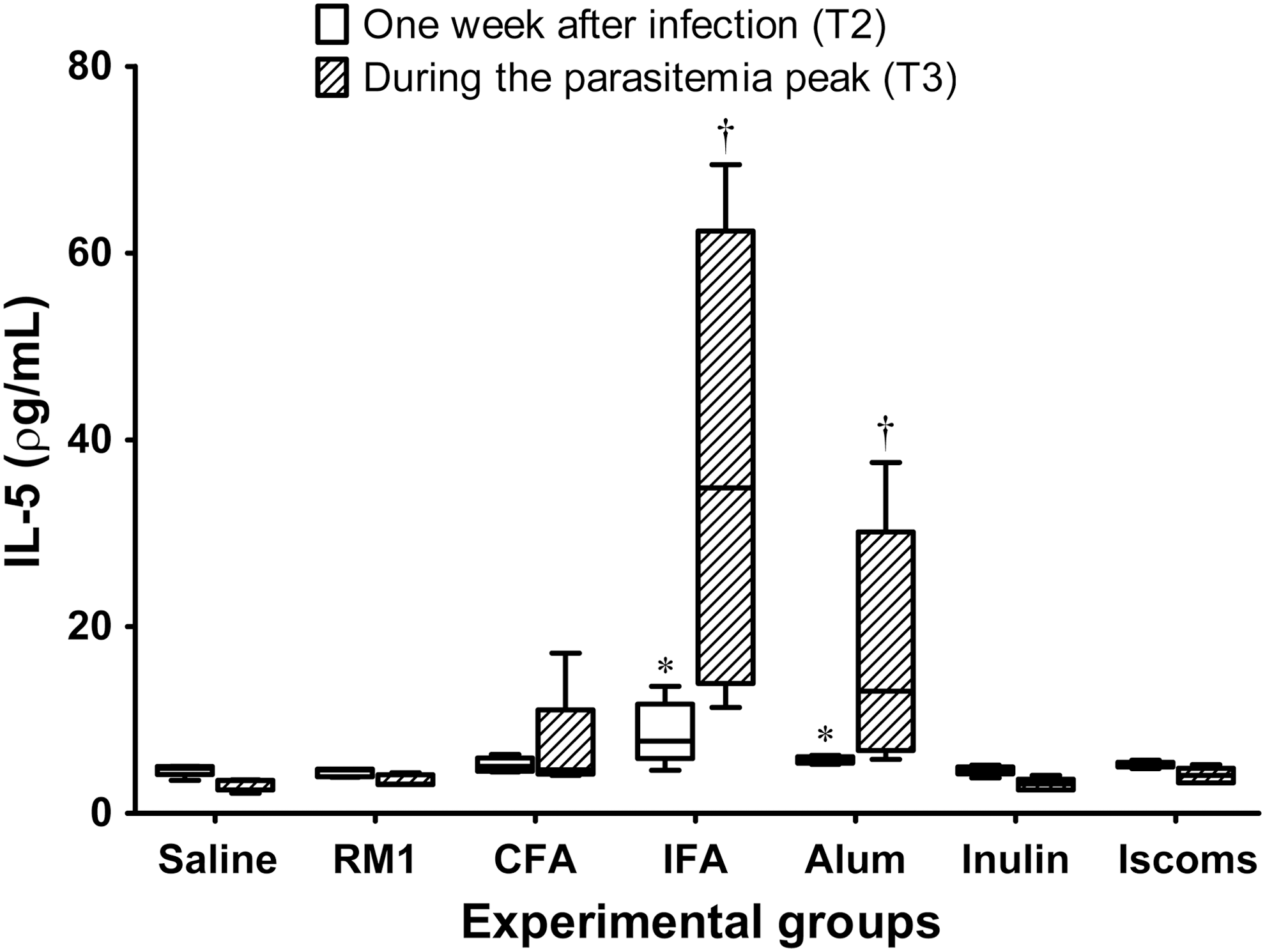
Plasma levels of IL-5 (pg/mL) from immunized animals with different adjuvants and challenged with T. cruzi Romildo strain. The graph illustrates only quantifications accomplished 1 week after infection (T2) and during the parasitemia peak (T3) for best viewing results. Nonparametric analysis was carried out for group comparisons by the Kruskal–Wallis test, followed by Dunn's post hoc tests. *p < 0.05 in comparison with saline, RM1, and Inulin groups 1 week after infection (T2). † p < 0.05 in comparison with saline, RM1, Inulin, and ISCOMs groups during the peak of parasitemia (T3).

Plasma levels of IFN-γ (pg/mL) from immunized animals with different adjuvants and challenged with T. cruzi Romildo strain. The graph illustrates only quantifications accomplished 1 week after infection (T2) and during the parasitemia peak (T3) for best viewing results. Nonparametric analysis was carried out for group comparisons by the Kruskal–Wallis test, followed by Dunn's post hoc tests. *p < 0.05 in comparison with saline, IFA, and Alum groups 1 week after infection (T2). † p < 0.05 in comparison with RM1, CFA, Inulin, and ISCOMs groups during the parasitemia peak (T3).
Discussion
The results of this study demonstrated that Th1/Th2 adjuvants have the capacity to direct the activation of the host immune response to a resistant and susceptible standard using an avirulent strain of trypanosomatid that is highly similar to T. cruzi as an immunogen.
The Th1 (CFA) or Th1/Th2 adjuvants (Inulin and ISCOMs) caused early production of IFN-γ, which increased 1 week after infection and was sufficient to act for the parasitemia control. The production of IFN-γ was slower in nonimmunized controls and in animals receiving Th2 adjuvants such as IFA and Alum and was observed after the parasitemia peak. Surprisingly, at this time, IFN-γ levels were increased in these animals, accompanied by high concentrations of IL-5, compared with other adjuvants. Although the IFN-γ represents a key cytokine of Th1 responses, animals that received standard Th2 adjuvants in the presence of T. cruzi marinkellei showed high levels of this cytokine precisely at the peak of parasitemia, in which the high count of blood trypomastigotes might contribute to immune activation of Th1 responses, characteristic of infection with T. cruzi.
Cytokines stimulated in response to infection, such as the case of IFN-γ, play an important role in the regulation of the host immune system, which indicates they are involved in both the resistance to T. cruzi and in the mechanisms related to the immunopathology of Chagas disease (Brener and Gazzinelli 1997, Michailowsky et al. 2001).
The antagonistic effect of IFN-γ in the resistance or susceptibility to infection indicates that it is a key cytokine in T. cruzi infection. During the acute phase, treated patients had higher levels of IFN-γ compared with untreated individuals. Conversely, during the chronic phase, high levels of IFN-γ were found in patients with cardiac or digestive manifestations compared with patients who had the indeterminate form (Dutra et al. 2005, Ribeiro et al. 2008). This duality of IFN-γ actions appears to have a temporary effect in which cytokine production in the acute phase of the infection reflects decreased susceptibility to the disease, whereas high production of IFN-γ during a chronic immune response results in the development of tissue lesions (Bahia-Oliveira et al. 1998). Although only the acute phase of infection was examined in the present study, animals presenting early peaks in IFN-γ levels (observed before the parasitemia peak) were able to control parasite replication more efficiently.
By stimulating a Th2 response, IFA and Alum were unable to protect mice from lethal infection. Conversely, Inulin, which stimulates a Th1/Th2 response, reduced parasitemia, but was not sufficient to increase the survival rate. Inulin is a hydrophilic adjuvant and, unlike hydrophobic adjuvants, does not have the deposit effect that delays the antigen liberation, resulting in a postponed immune response (Petrovsky and Aguilar 2004). The use of ISCOM as an adjuvant was able to reduce parasitemia and increase the survival rate. However, animals had increased muscle tissue inflammation, which may have contributed to a possible imbalance in the host immune system during the infection by T. cruzi.
Host immunomodulation is therefore central to the development of a vaccine against T. cruzi (Cunha-Neto et al. 2009, Araujo et al. 2011). The presence of T cells that coexpress IFN-γ and IL-10, for example, may be evidence of the long-term presence of T. cruzi in the host combined with reduced tissue damage resulting from the regulation and suppression of the immune response against the parasite (Flores-Garcia et al. 2011). Basso (2013) reported that the IFN-γ/IL-10 ratio is higher in animals that had not been previously immunized, which suggests that excessive proinflammatory cytokines may indicate poor prognosis (Basso 2013). It is well established that the presence of anti-inflammatory cytokines is essential for controlling the tissue damage resulting from an excessive immune response where the constant activity of IFN-γ and TNF-α may lead to tissue damage, cardiac dysfunction, and death (Dutra et al. 2014, Sousa et al. 2014). In the present study, we did not observe detectable levels of IL-10 in the plasma, although this cytokine may be locally produced by different subsets of cells present in the inflammatory microenvironment in animals with reduced tissue damage. Further studies are necessary to quantify the cytokine levels in the tissue sections.
Accordingly, the development of new adjuvants is necessary for the identification of its mechanism of action within the host immune system. With the increase in immune stimulation by adjuvants, the decision to induce local or systemic reactions must be considered so that it is possible to obtain a maximal immune response with minimal adverse effects (Lindblad 2007).
In this context, it is also important to consider the possible autoimmune mechanisms in the pathogenesis of Chagas disease, which can be explained by bystander activation, molecular mimicry, polyclonal activation, cryptic epitopes, and epitope spreading (Machado et al. 2012a). Among these mechanisms, the evidence that some T. cruzi epitopes share peptide sequences with cardiac myosin reinforces the need for the identification of protective antigens that do not increase inflammation for the development of a vaccine against Chagas disease (Miyahira 2008, Arce-Fonseca et al. 2015). Antigenic similarities between different trypanosomatids, demonstrated by their strong immunogenicity, support the use of these strains as possible components of a vaccine for Chagas disease.
It is important to highlight that in all of the evaluated parameters, animals immunized with the RM1 strain without adjuvant presented the best results, such as substantial reduction in parasitemia and increase in survival. Furthermore, this immunization strategy activated few inflammatory processes with early and controlled production of IFN-γ and lower levels of IL-5. These findings indicate that the RM1 strain may comprise antigens that are potent activators of the immune response. We were expected to get better protective immunity mediated by Th1 adjuvants, which are able to induce the production of proinflammatory cytokines, such as IFN-γ and TNF-a. However, the additional stimulus provided by adjuvants may have negatively affected the immune balance.
For this reason, future studies are needed to identify conserved antigens in this strain, which are important for the development of prophylactic and therapeutic vaccines, and to demonstrate how these trypanosomatids can induce CD8+ T lymphocyte activation, which are identified as the main cells in immunoprotection against T. cruzi (Dos Santos Virgilio et al. 2014). Furthermore, the use of other avirulent T. cruzi strains or the search for the antigenic characterization of trypanosomatids isolated from bats deserves particular attention because they make a connection between the sylvatic and domestic cycles of Chagas disease, which may contribute to the antigenic diversity in trypanosomatids and the generation of a protective and balanced immune response in the absence of adjuvants.
Finally, immune evasion of T. cruzi makes difficult the development of a vaccine that confers sterile immunity (Cazorla et al. 2009), especially in view of their different infective stages (Rodriguez-Morales et al. 2015). However, a vaccine that is able to modulate the immune response would still be useful to avoid imbalance in the host–parasite dynamics, particularly when this relationship drives to the progression of the symptomatic disease.
Footnotes
Acknowledgments
This work was supported by grants from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES—Edital 32/2010, Programa de Incentivo à Pesquisa em Parasitologia), Fundação de Amparo à Pesquisa de Minas Gerais (FAPEMIG—CBB-APQ 2308/10), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq—142614/2007-6).
The authors thank Professor Cristina Ribeiro de Barros Cardoso, from the School of Pharmaceutical Sciences at Ribeirão Preto/USP, Brazil, for her scientific contribution in immunoassays and Professor João Santana da Silva for the permission to use the Laboratory of Immunoparasitology facilities in the School of Medicine at Ribeirão Preto, University of São Paulo (FMRP-USP). The authors also thank Luciano Henrique Paiva for the technical assistance.
Author Disclosure Statement
No competing financial interests exist.