
Abstract
Batai virus (BATV) is an arthropod-borne single-stranded RNA virus belonging to the genus Orthobunyavirus of the family Bunyaviridae that is primarily transmitted by mosquitoes. Methods for detecting BATV are currently limited to serological surveillance, virus isolation, and conventional reverse transcription–polymerase chain reaction (RT-PCR) assay. In this study, we sought to develop a BATV detection assay that needs no specialized equipment and is highly specific, sensitive, and simple. We first developed and optimized a reverse transcription loop-mediated isothermal amplification (RT-LAMP) for rapid detection of BATV that uses two pairs of primers to amplify a conserved region of the BATV M gene. The optimal reaction conditions for this RT-LAMP BATV detection assay were 40 min at 65°C. The amplification products could be visualized directly for color changes. This RT-LAMP method has a detection limit of 2.86 copies/μL and a sensitivity that was approximately 10- and 100-fold greater than real-time and conventional RT-PCR, respectively. RT-LAMP for BATV detection showed no cross-reactivity with other viruses and its sensitivity was validated with cattle blood and mosquito specimens. Our results suggest that this RT-LAMP method was simpler and faster than conventional RT-PCR or real-time RT-PCR. Moreover, RT-LAMP represents a potential tool to test for BATV in clinical and mosquito samples, especially in rural areas of China. This method also shows promise as a diagnostic tool due to its rapid and sensitive detection without the need for sophisticated equipment or complicated protocols.
Introduction
B
Two BATV strains (YN92-4 and NM/12) were recently isolated from two different hosts (Anopheles Philippines and cattle) in China (Liu et al. 2014). The BATV genome consists of three single-stranded RNA segments of negative polarity that are designated S (small), M (medium), and L (large). The S RNA segment encodes the nucleocapsid protein (N) and a nonstructural protein (NSs). The M segment codes for a glycoprotein precursor, which is cleaved by host proteases into a nonstructural protein (NSm) and two virion surface glycoproteins, Gn and Gc. The L segment encodes the RNA-dependent RNA polymerase (RdRp). Orthobunyavirus species can increase their genetic diversity by reassortment of these genome segments during mixed infections (Yanase et al. 2006). For example, a genetic reassortment of the BATV M segment and the S and L segments from the Bunyamwera virus generated the Ngari virus (Groseth et al. 2012).
Early detection and diagnosis might help lower the spread of BATV and minimize disease-related economic losses. To date, there are few studies that investigated the distribution and prevalence of BATV using viral isolation and gene amplification-based molecular techniques such as reverse transcription–polymerase chain reaction (RT-PCR). Although conventional RT-PCR is the preferred method for viral diagnoses of infected host tissue and fluid samples, in rural areas where BATV transmission is prevalent, there is a lack of RT-PCR equipment and technical support. In addition, the low copy numbers of viral RNA in the blood of BATV-infected hosts also limit RT-PCR-based BATV detection.
Rapid, sensitive, and simple diagnostic methods to detect BATV are critical for more effective disease control. Reverse transcription loop-mediated isothermal amplification (RT-LAMP) is a relatively new nucleic acid amplification method that can amplify DNA rapidly and efficiently with high specificity under isothermal conditions (Notomi et al. 2000). This method relies on autocycling strand displacement DNA synthesis performed by the Bst DNA polymerase large fragment. The most important advantage of the LAMP method is its ability to amplify specific target DNA sequences with high specificity and sensitivity within 1 h or less under isothermal conditions at a temperature range of 60–65°C. Moreover, no thermal cycler and/or specialized laboratory facilities are needed. The higher amplification efficiency of the LAMP method enables simple visual observation of amplification using intercalating dyes such as SYBR Green I with UV illumination (Mori et al. 2001, Nagamine et al. 2002) or digital imaging after agarose gel electrophoresis. The RT-LAMP method has been developed to detect a variety of viruses, including foot-and-mouth disease virus (Yamazaki et al. 2013), dengue virus (Parida et al. 2005), rubella viruses (Abo et al. 2014), and West Nile virus (Parida et al. 2004), but has not yet been applied to detect BATV. In this study, we sought to develop the LAMP method to detect BATV in mosquitoes and blood samples from cattle. Our results suggest that the RT-LAMP method is more sensitive than RT-PCR and would be a potential tool to test for the presence of BATV in mosquitoes and clinical samples from rural areas of China.
Materials and Methods
Virus strains
Based on its previous isolation from cattle, the BATV NM/12 strain belonging to the MM2222 lineages was selected as a positive standard (Liu et al. 2014). Porcine reproductive and respiratory syndrome virus (PRRSV, CC-1; GenBank: EF153486), bovine viral diarrhea virus (BVDV, HLJ-10; GenBank: JF714967), Akabane disease virus (AKAV, NM/12; sequence has not been deposited in the GenBank database), and Japanese encephalitis virus (JEV, YN0901; GenBank: JQ086762) were isolated and identified by our laboratory and stored at −80°C until use.
Sample collection
Adult female mosquitoes from cattle sheds and hog pens were trapped at dusk using ultraviolet lamps as attractants. The collected mosquitoes were distributed among 135 tubes (100 mosquitoes/tube) and identified and sorted according to species; only Culex females were used for these experiments. Cattle blood samples (126 total), including 3 positive samples and 123 negative serum samples, were obtained from different farms in Inner Mongolia, China (110°46′–112°10′ N, 40°51′–41°8′ E), and analyzed using a PCR method that was designed and previously reported (Yanase et al. 2006).
RNA extraction from the NM/12 strains and reverse transcription to cDNA
Working virus stocks of NM/12 strain BTV were propagated in Vero cell cultures, and culture supernatants containing the virus were harvested at 72 h post-infection. The cellular debris was removed from 0.5 mL culture supernatant that contained 107 plaque-forming units by centrifugation before the viral RNA was extracted using the QIAamp Viral RNA Mini Kit (Qiagen) in accordance with the manufacturer's protocol. To evaluate the assay detection limit, the amount of BATV RNA was determined by spectrophotometry and converted to molecular copies using a previously published formula (Krieg 1990).
Tenfold serially diluted BATV RNA (2.86 × 105–2.86 × 10−1 copies/μL) was used as a template for reverse transcription. Briefly, 9.0 μL diethyl pyrocarbonate (DEPC)-treated H2O, 1.0 μL of 50 ng/μL genotype-specific reverse primer R1 (Table 1) (Shanghai Sangon Biological Engineering Technology and Service), and 5.0 μL total RNA were mixed and incubated at 75°C for 5 min and chilled on ice for at least 5 min. The following reagents were then added to the reactions: 9.0 μL DEPC H2O, 1.0 μL 200 U/μL M-MLV RT (Promega), 1.0 μL 40 U/μL ribonuclease inhibitor (Promega), 6.0 μL 5× RT buffer, and 2.0 μL 10 mM dNTP mix (TaKaRa). The reaction mixture was incubated at 42°C for 50 min and 75°C for 5 min, and the cDNA was used as a template for the RT-LAMP and PCR assays (Liu et al. 2012).
Nucleotide positions: amplicons were designed to correspond with the conserved regions of BATV NM/12 strain. Primers for conventional RT-PCR and RT-LAMP amplification of the BATV M genes. Primers for real-time RT-PCR amplification of the BATV S genes.
BATV, Batai virus; RT-LAMP, reverse transcription loop-mediated isothermal amplification; RT-PCR, reverse transcription–polymerase chain reaction.
Primer design for RT-LAMP and RT-PCR of BATV
We used the Primer Explorer version 3 software for RT-LAMP assay to design two pairs of primers with sequences that were based on alignment analysis of genomic sequences of 15 complete Batai virus M segments deposited in GenBank. Amplicons were designed in conserved regions (Fig. 1). Each set of primers included an outer pair (F3, B3) and an inner pair (FIP, BIP). The primers for conventional RT-PCR amplification of the BATV M genes were M1 and M2. The resultant BAT FP, BAT P, and BAT RP primers were used for real-time RT-PCR amplification of the BATV S genes (Table 1). Primers were synthesized by the Shanghai Sangon Biological Engineering Technology and Service (Shanghai, China).

Location of loop-mediated isothermal amplification primers in the BATV M gene. The primers included two outer primers (forward primer, F3, and reverse primer, B3) and two inner primers (forward inner primer, FIP, and reverse inner primer, BIP). Color images available online at
RT-LAMP assay optimization
The RT-LAMP reaction mixture contained 1.0 μL each of 5.0 μM F3 and B3 primers, 1.0 μL each of 40 μM BIP and FIP primers, 2.5 μL 10× ThermoPol reaction buffer, 1.0 μL 8 U/μL Bst DNA polymerase (New England Biolabs), 2.5 μL 10 mM dNTP mix (TaKaRa), 4.0 μL 5 M betaine (Sigma-Aldrich), 5.0 μL 30 mM MgSO4 (Invitrogen), 2.0 μL target cDNA, and 4.0 μL nuclease-free water (Liu et al. 2012). The mixtures were incubated at 59, 60, 61, 62, 63, 64, or 65°C for 60 min to determine the optimal reaction temperature. Subsequently, the optimal reaction time was determined by performing RT-LAMP at 65°C for 10, 20, 30, 40, 50, or 60 min. The reactions were terminated by heat inactivation at 80°C for 10 min. Sterile water was used in place of template cDNA as a negative control. RT-LAMP products (10 μL) were resolved by 2.5% agarose gel electrophoresis with ethidium bromide staining visualized by ultraviolet light. In an alternate protocol, the products were observed directly with the naked eye by adding 1.0 μL SYBR Green I (10,000×; Invitrogen) to the reaction mix. The solution turned green in the presence of RT-LAMP amplification products, but remained orange in the absence of the amplicon. The replicates were performed at least thrice.
Conventional RT-PCR of the BATV M gene
Conventional RT-PCR mixtures contained 1.0 μL 5 U/μL ExTaq polymerase (TaKaRa), 0.5 μL 10 mM dNTP mix, 0.5 μL each of M1 and M2 primers (10 μM), 2.0 μL ExTaq buffer, 2.0 μL cDNA, and 13.5 μL nuclease-free water. The PCR was performed using a TC-512 instrument (Techne) with the following reaction conditions: one cycle at 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 56°C for 30 s, and 72°C for 30 s, and a final extension cycle at 72°C for 10 min. The products (10 μL) were analyzed by 1.5% agarose gel electrophoresis with ethidium bromide staining visualized by UV light. The expected product size was 557 bp. The replicates were performed at least thrice.
Determination of real-time PCR assay detection limit
Real-time RT-PCR was performed to amplify BATV as described in a previous report. (Weidmann et al. 2003). RT-PCR thermal cycling programs were run on a Roche LightCycler 96 Real-Time PCR System.
Specificity of RT-LAMP assay for BATV
To analyze the specificity of the established RT-LAMP assay, BATV (NM/12; GenBank: KJ187039), PRRSV (CC-1; GenBank: EF153486), BVDV (HLJ-10; GenBank: JF714967), AKAV (NM/12; not deposited in the GenBank database), and JEV (YN0901; GenBank: JQ086762), which cause similar diseases in cattle/pigs, were used as RT-LAMP templates. The reaction was performed at 65°C for 40 min at least thrice. We aligned M segments of hantavirus (HV), snowshoe hare virus (SSHV), Jamestown Canyon virus (JCV), Oropouche virus (OROV), La Crosse virus (LACV), Inkoo virus (INKV), and Tahyna virus (TAHV), together with the primer sequences. Amplicons were aligned using Megalign software (DNAstar; Lasergene) and BLAST in Genbank.
Validation of BATV RT-LAMP, RT-PCR, and real-time RT-PCR assays with swine blood samples and mosquito samples
We compared the RT-PCR, real-time RT-PCR, and LAMP results with previous serum samples and mosquitoes; replicates were performed at least thrice. The samples (true-positive or false-positive) were determined from the results of isolation and identification of BATV.
Virus isolation from cattle blood and mosquito samples and identification of the virus using RT-PCR
The 126 cattle blood samples, 3 suspected positive samples, and 123 negative serum samples were separated into plasma and blood cells. The blood cells were washed with phosphate-buffered saline (PBS) and the plasma and blood cells were resuspended in PBS and stored at −80°C until virus isolation. A total of 135 tubes containing female Culex tritaeniorhynchus mosquitoes were used for these experiments. Viral suspensions from blood and C. tritaeniorhynchus females (20 μL) were propagated for 72 h in Vero cells in complete minimum essential medium supplemented with 5% fetal calf serum and 1% penicillin (10,000 U/mL)/streptomycin (10,000 μg/mL) at 37°C in a 5% CO2 environment and the cytopathogenic effect was observed. Viral RNA was reverse transcribed and amplified according to the manufacturer's instructions and the method described in a previous report (Yanase et al. 2006). RT-PCR products (10 μL) from all examined samples with the expected size corresponding to the partial BATV M gene were identified in 1.5% agarose gels. To achieve high-quality consensus sequences and avoid laboratory PCR artifacts, the entire viral genome was sequenced at least thrice, with one sequence obtained from an uncloned PCR product. The amplified fragment was cloned into the pMD18-T vector and confirmed by sequencing.
Results
RT-LAMP amplification of the BATV M gene
Agarose gel electrophoresis of DNA products amplified by RT-LAMP showed a characteristic laddering pattern with the resolution of multiple bands, indicating that the final RT-LAMP products were mixtures of stem-loop DNAs of various stem lengths (Fig. 2). In contrast, the negative control lacked this characteristic multiband laddering pattern.

RT-LAMP amplification of the BAT M gene. Lanes: M, marker; 1, RT-LAMP product; 2, negative control. RT-LAMP, reverse transcription loop-mediated isothermal amplification.
Optimization of BATV RT-LAMP
The optimal reaction temperature and time of BATV RT-LAMP assay were next investigated. RT-LAMP reactions at 65°C produced stronger band intensities than those seen for reactions at other temperatures (Fig. 3). The RT-LAMP assay reactions were then run for varying lengths of time, which showed that RT-LAMP at 65°C with a 40-min reaction time yielded appropriate apparent band intensities (Fig. 4). Therefore, the optimal reaction conditions of the RT-LAMP assay for detecting BATV were 65°C for 40 min.
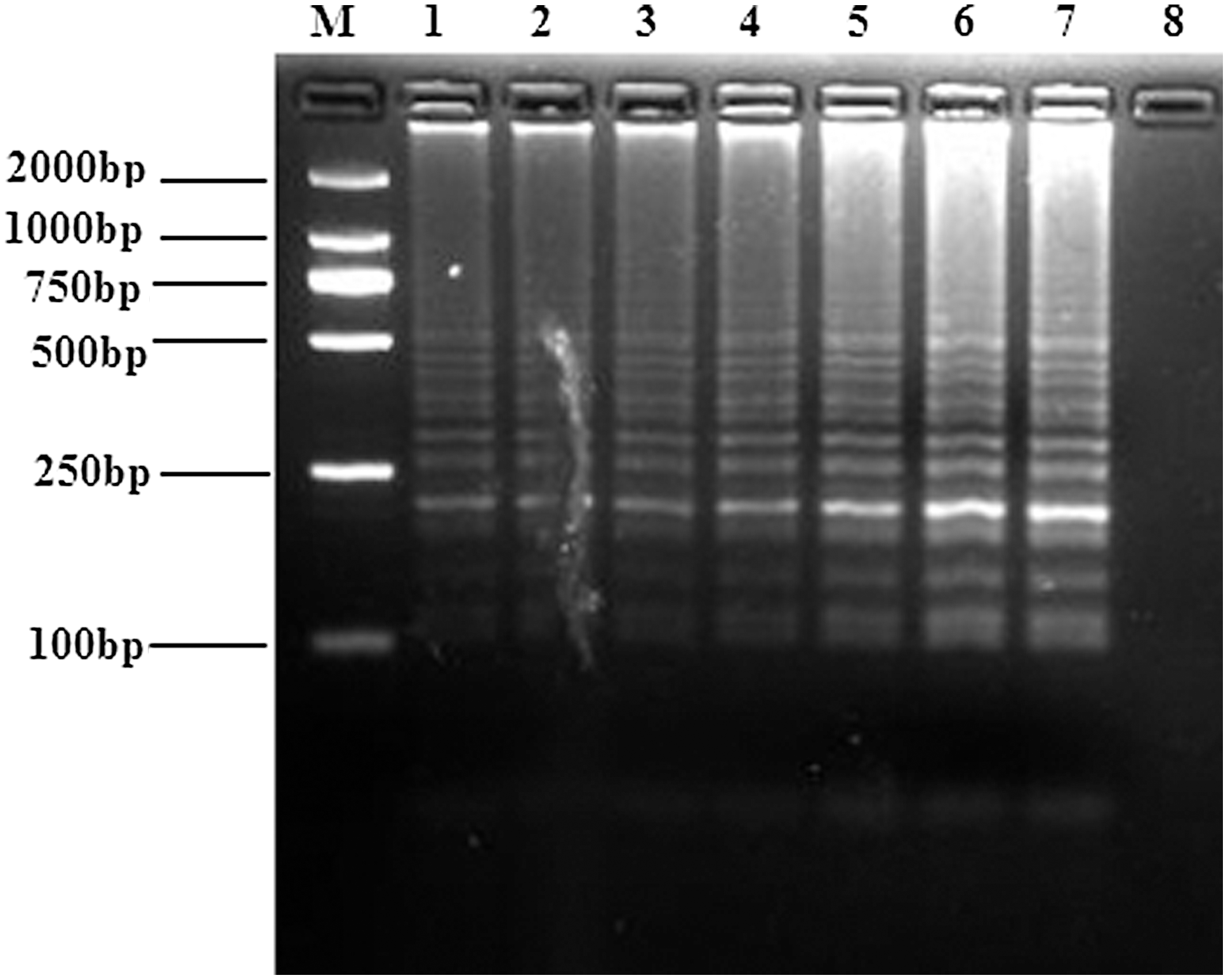
Optimization of the temperature for RT-LAMP detection of the BATV M gene. RT-LAMP reactions were performed at 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, or 65°C for 60 min. The reaction products were resolved on an agarose gel, and the results are shown in lanes 1–7. Lanes: M, DNA marker; 8, negative control.
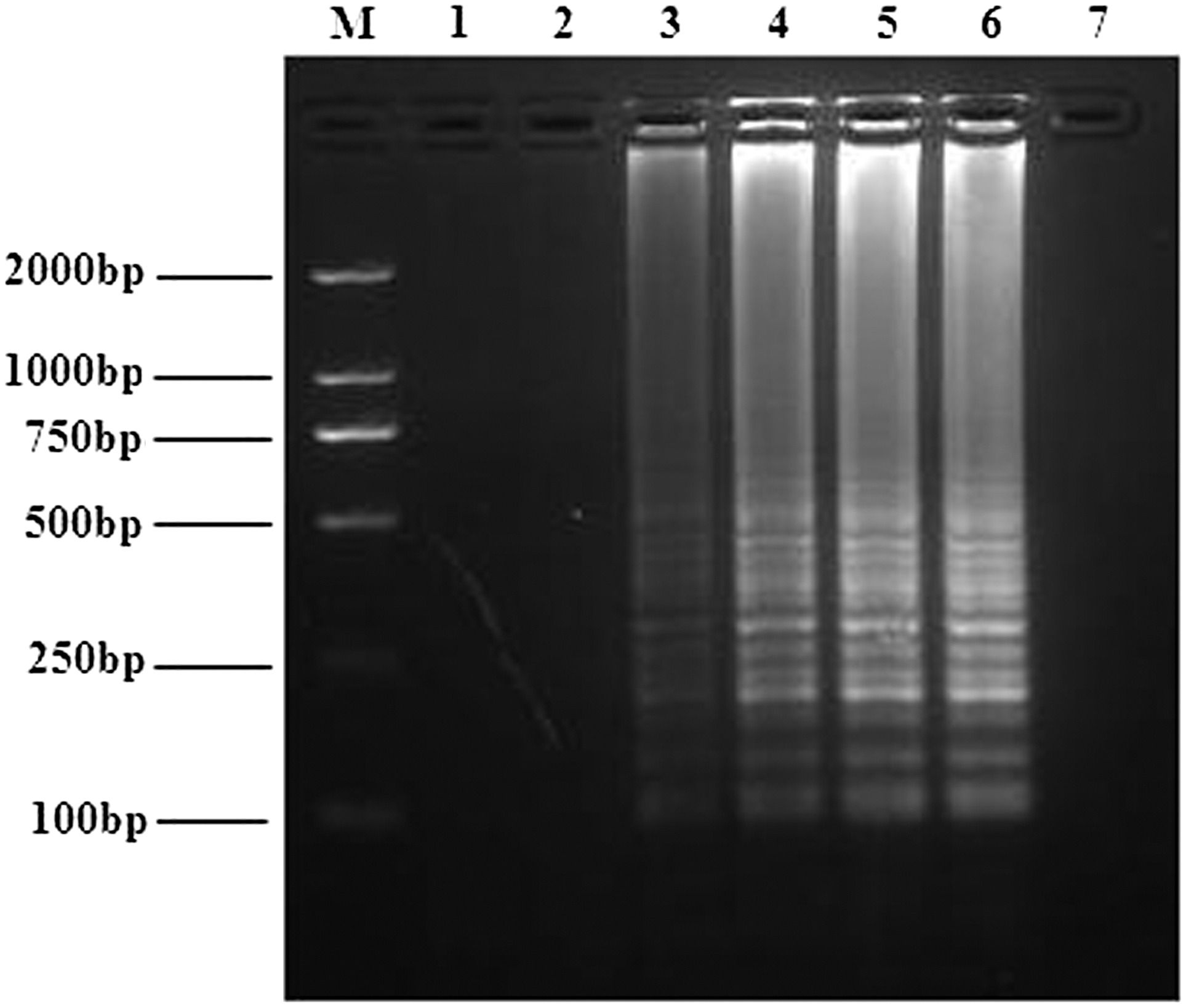
Optimization of the reaction time for RT-LAMP detection of the BATV M gene. RT-LAMP reactions were performed at 65°C for 10, 20, 30, 40, 50 or 60 min. The reaction results for the six different reaction time periods are shown in lanes 1–6, respectively. Lanes: M, DNA marker; 7, negative control.
BATV detection limits of the RT-LAMP assay, conventional RT-PCR, and real-time RT-PCR
We next compared the detection limits of the RT-LAMP assay, real-time RT-PCR, and conventional RT-PCR for BATV detection. Tenfold serial dilutions of BATV RNA (from 105 to 10−1 copies/μL) were used to determine assay sensitivity wherein positive reaction mixtures showed a characteristic ladder-like pattern. Samples containing 2.86 copies/μL were detected 100% of the time across three replicates, while more dilute samples (presumably 0.286 copies/μL) were not detected (Fig. 5A). Moreover, visual inspection after SYBR Green I staining revealed stronger intensities of the amplification products from the RT-LAMP assay than from conventional RT-PCR and real-time RT-PCR (Figs. 5B–D and 6A, B). Visualization with UV transillumination revealed detection limits of 2.86 BATV copies/μL by RT-LAMP, which was 100- and 10-fold more sensitive than conventional RT-PCR and real-time RT-PCR assay, respectively (Table 2).
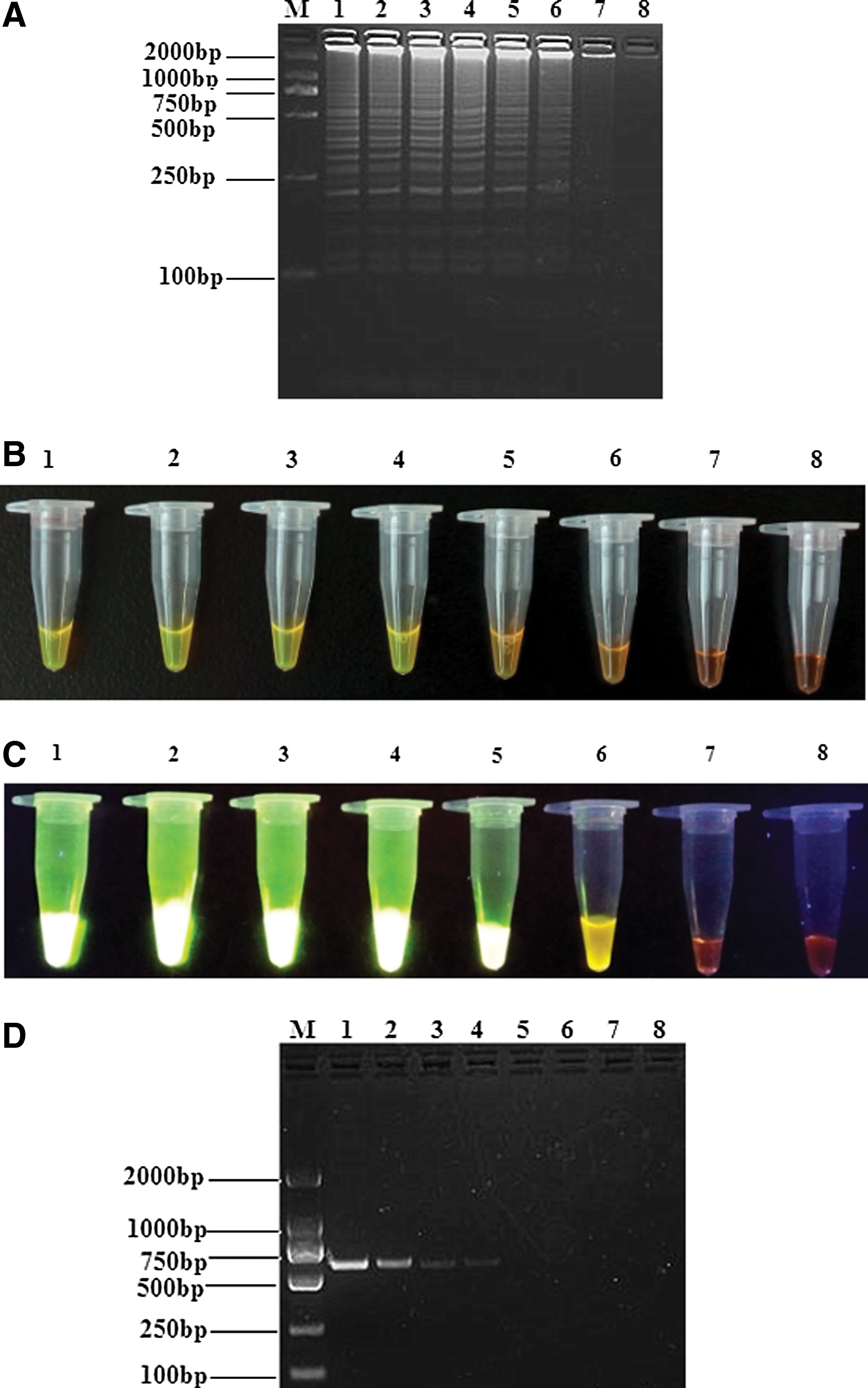
Comparison of RT-LAMP and RT-PCR assay sensitivity for BATV detection.
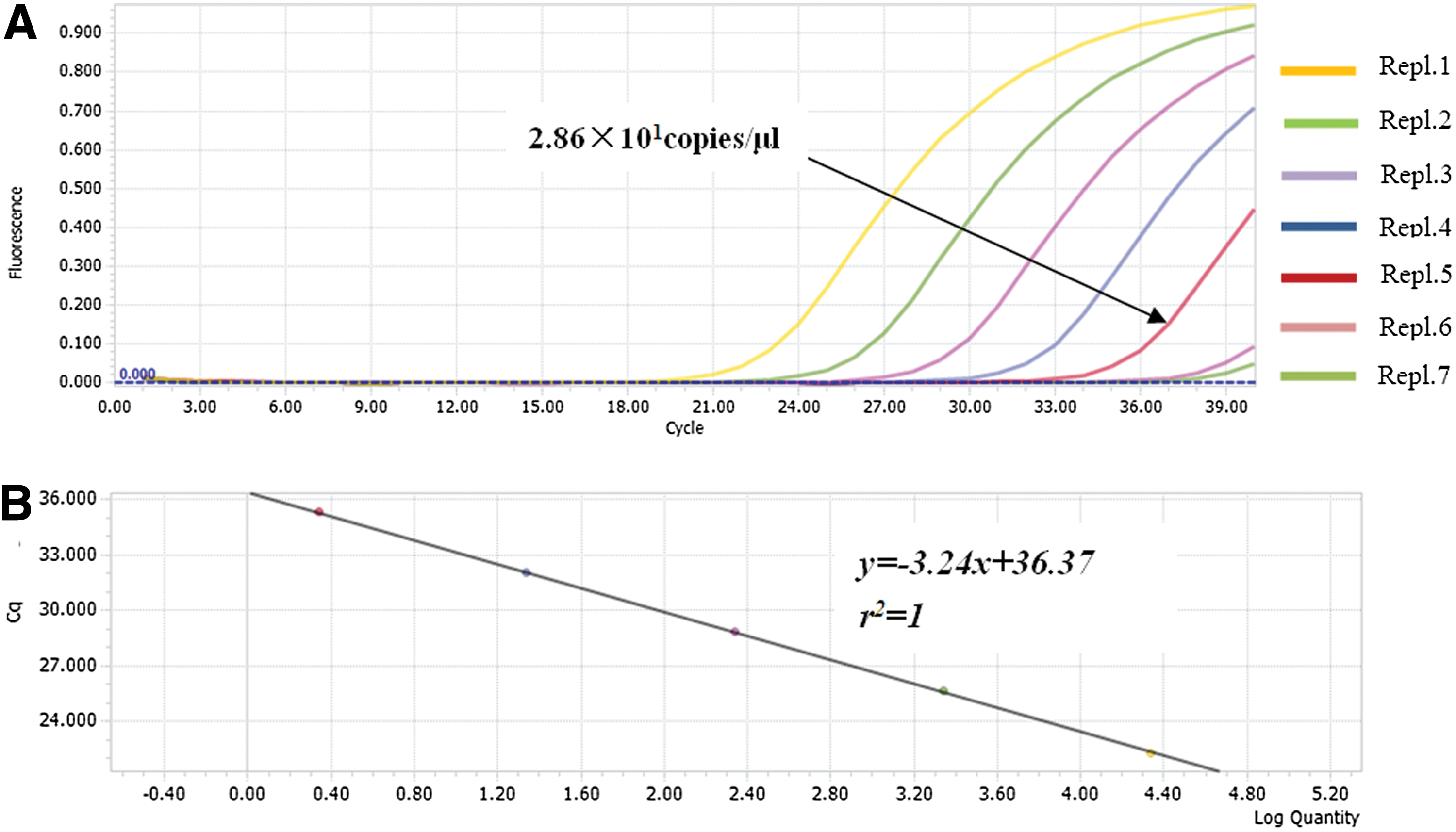
Detection sensitivity of the real-time RT-PCR assay for BATV RNA.
+, positive reaction; –, negative reaction.
RT-LAMP assay specificity
No amplification product was observed for reaction mixtures that contained cDNA templates from PRRSV (CC-1), BVDV (HLJ-10), AKAV (NM/12), and JEV (YN0801), suggesting that RT-LAMP is highly specific for BATV (Fig. 7). Alignment of M segments from HV, SSHV, JCV, OROV, LACV, INKV, and TAHV strains showed no sequence similarity with the BATV primers used here, indicating that there would be a low possibility of detecting other Orthobunyavirus genus members using BATV-specific primers with RT-LAMP.
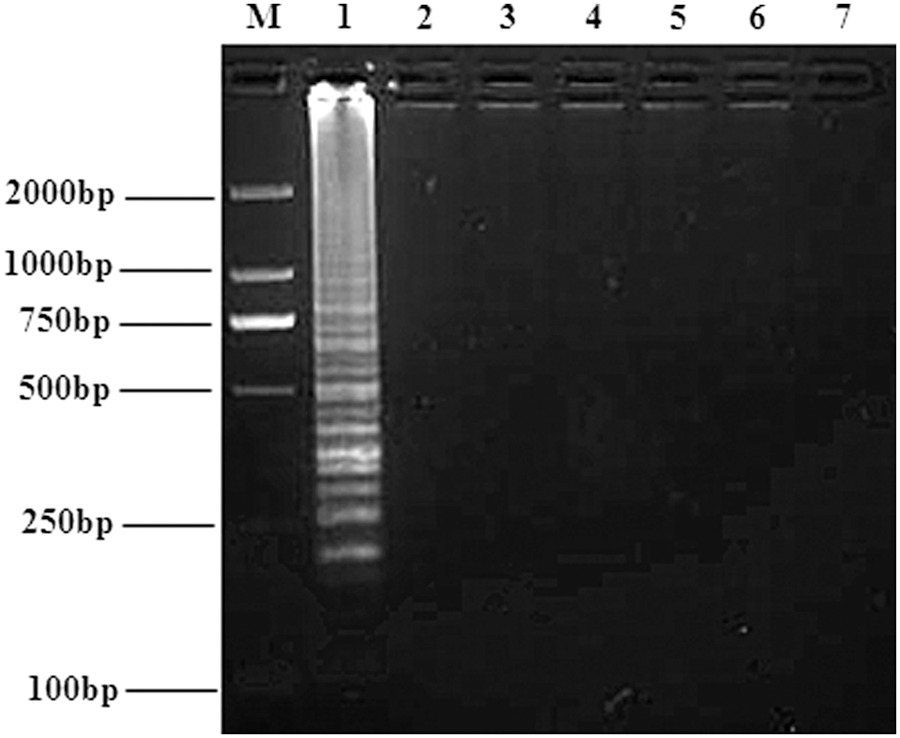
Specificity of the RT-LAMP assay for detecting the BATV M gene. BATV and other selected viruses were used as templates and subjected to RT-LAMP performed at 65°C for 40 min. Lanes: M, DNA marker; 1–5, RT-LAMP results from templates BATV (NM/12), PRRSV (CC-1), BVDV (HLJ-10), AKAV (NM/12), and JEV (YN0801), respectively; 6, negative control. AKAV, Akabane disease virus; BVDV, bovine viral diarrhea virus; JEV, Japanese encephalitis virus; PRRSV, Porcine reproductive and respiratory syndrome virus.
Validation of RT-LAMP, conventional RT-PCR, and real-time RT-PCR methods to detect BATV in mosquito samples and cattle serum samples
All 121 negative serum samples were also negative by all tests, thus ruling out the possibility of false positives. Furthermore, none of the RT-PCR or real-time RT-PCR-positive samples was missed by RT-LAMP, which demonstrated much higher sensitivity than either conventional RT-PCR or real-time RT-PCR (Tables 3 and 4). Analysis of mosquito RNA samples by RT-PCR and real-time RT-PCR showed two false results and one false result, respectively, for BATV-negative samples. Results of BATV virus isolation and identification in cattle blood samples and mosquito samples were consistent with those for RT-LAMP. The sequence analysis of the positive samples revealed the highest sequence identity with BATV, thereby confirming the higher sensitivity of the RT-LAMP assay.
RT-LAMP, reverse transcription loop-mediated isothermal amplification.
Discussion
Serological surveillance and virus isolation show that BATV is widely distributed worldwide. However, the application of conventional assays, such as serological surveillance and virus isolation, is limited by requirements for laboratory-based operations and skilled technicians, and the sensitivity of these assays is much lower than molecular diagnostic techniques. Therefore, to identify BATV infection at a molecular level, genome-based RT-PCR and real-time RT-PCR methods have been developed. These methods also require equipment and technical support that is often lacking in rural areas where BATV is prevalent, which in turn limits the use of RT-PCR and real-time RT-PCR for BATV detection.
In contrast, the RT-LAMP assay is an isothermal amplification technique that can amplify DNA at constant temperatures. Thus, LAMP can be an effective diagnostic test for BATV in developing countries as no sophisticated equipment or skilled personnel are needed for this assay. LAMP is also a cost-effective testing approach (Parida et al. 2008; Dhama et al. 2014).
In this study, we first developed an RT-LAMP method for BATV detection and found that the optimal reaction conditions for BATV M gene amplification were 65°C and 40 min. The reaction products exhibited characteristic ladder-like patterns following agarose gel electrophoresis and were detectable by naked eye inspection with SYBR Green I staining under normal light or UV transillumination. The detection limit of RT-LAMP was 10- and 100-fold higher than real-time RT-PCR and conventional RT-PCR, respectively. This outcome is consistent with several reports, which showed that LAMP exhibits higher sensitivity (<10 copies) compared with regular conventional RT-PCR and real-time RT-PCR (Weidmann et al. 2003, Teoh et al. 2013, Shirato et al. 2014). The mosquitoes and cattle serum samples that were positive by conventional RT-PCR and real-time RT-PCR were also positive using the RT-LAMP assay. Meanwhile, RT-PCR for BATV produced four and two false outcomes from mosquito and cattle serum samples, respectively, and real-time RT-PCR showed one false outcome for both sample types. Thus, the RT-LAMP assay has higher sensitivity than conventional RT-PCR and real-time RT-PCR. In addition, the BATV RT-LAMP assay showed good specificity as no positive results were found using RNA from other viruses such as PRRSV (CC-1), BVDV (HLJ-10), AKAV (NM/12), and JEV (YN0801). Sequence alignments between the M segment BATV sequences and other Orthobunyavirus genus members confirmed that RT-LAMP used with these primers would not detect other orthobunyaviruses. Moreover, the RT-LAMP assay showed good repeatability across three trials. Further studies will be needed to determine how the RT-LAMP assay performs with other known BATV strains since only one Chinese BATV strain was tested in the current study.
In summary, we describe the LAMP method that is rapid, sensitive, and specific for the detection of BATV in mosquito and cattle serum samples. As this RT-LAMP method is easy to establish and requires no additional equipment, RT-LAMP should serve as a promising detection tool for early BATV diagnosis in rural clinics and field situations where resources are limited.
Footnotes
Acknowledgments
The authors wish to express their sincere gratitude to Ning-Yi Jin at the Institute of Military Veterinary, Academy of Military Medical Sciences, for excellent technical assistance. This work was supported by the Technology and Development Program of Jilin Province (20150520127JH).
Author Disclosure Statement
No competing financial interests exist.