
Abstract
Members from Rickettsiales such as Anaplasma phagocytophilum, Ehrlichia chaffeensis, and some spotted fever group (SFG) Rickettsiae are important tick-borne pathogens. One hundred goats and sheep from southeastern China were examined for the presence of Anaplasma, E. chaffeensis, and SFG Rickettsiae by PCR. A. phagocytophilum, Anaplasma bovis, and Anaplasma centrale were detected in 15, 49, and 16 samples, respectively. The A. phagocytophilum and A. centrale were highly homologous to strains from Japanese sika deer and Japanese cattle, respectively, whereas a diversity of A. bovis sequences were detected. New genetic variants of Anaplasma close to A. centrale have been revealed. No Ehrlichia was detected in this study. The presence of SFG Rickettsiae was determined in 26 samples. The coinfection with more than two pathogens tested in this study was as high as 29%. This study has molecularly characterized the circulation of Anaplasma and Rickettsiae in goats and sheep in southeastern China, which highlights the risk of contracting the pathogens upon tick exposure.
Introduction
R
The presence of Anaplasma, Ehrlichia, and SFG Rickettsia in ticks, animals, or humans has been reported in China (Wang and Walker 1987, Wen et al. 2003, Cao et al. 2006, Zhang et al. 2008, 2009, Dong et al. 2013). SFG Rickettsiae were previously reported to be prevalent mainly in western and northern China (Fan et al. 1987, Yu et al. 1993). In recent years, however, it has been found that they are also present in central and southern China (Li et al. 2009, Zou et al. 2011, Liang et al. 2012, Tian et al. 2012). Compared with tick or wild rodent hosts, less research has been performed on the detection of Anaplasma, Ehrlichia, or SFG Rickettsia in domestic animals in southeastern China.
Our previous study has described the presence of Theileria luwenshuni, a tick-borne hemoparasitic pathogen in goats from southeastern China (Ge et al. 2012). The present study further investigates the prevalence of Rickettsiales in the same sample sets as described previously (Ge et al. 2012) from southeastern China by the nested PCR amplification of specific genes and sequence analyses.
Materials and Methods
Collection of field samples
The study was carried out in 10 ranching farms from central Zhejiang province in southeastern China (Fig. 1). During July 2010, a total of 90 and 10 clinically healthy goats and sheep, respectively, from these farms were randomly selected by 10 samples from each farm (Table 1). Blood was drawn as previously described (Ge et al. 2012). This study was approved by the Animal Ethics Committee of Tongji University School of Medicine.

The collection sites of goat and sheep blood samples from Zhejiang province, China.
Ten sample animals from farm B-3 marked with * were sheep and 90 sample animals from the other nine farms were goats.
SFG, spotted fever group.
DNA extraction and PCR amplification
The genomic DNAs from the blood samples were extracted as previously described (Ge et al. 2012). To detect Anaplasma species, a nested PCR was carried out to amplify partial 16S rRNA genes. Primers EC12A and EC9 were used in the first PCR to amplify all Anaplasma spp. Based on our preliminary study, the three Anaplasma species of A. phagocytophilum, Anaplasma centrale, and Anaplasma bovis were selected for investigation. Species-specific primers for A. phagocytophilum, A. bovis, and A. centrale were used in the secondary PCR. A nested PCR with genus-specific primers was carried out to amplify the Anaplasma partial citrate synthase gene (gltA). To detect E. chaffeensis, a nested PCR was performed to amplify partial 16S rRNA genes. To detect SFG Rickettsia spp., a nested PCR was conducted to amplify a region of the rompB gene. All the primers used in this study are listed in Table 2. DNAs from A. phagocytophilum HZ strain, E. chaffeensis Arkansas strain, and Rickettsia heilongjiangensis 054 strain were used as positive controls. Ultrapure water was used as a negative control in all PCRs. PCR reactions was performed as previously described (Ge, et al. 2012), and cycle conditions were set as described in the references in (Table 2).
Cloning, sequencing, and sequence analysis
The PCR amplicons were gel purified and then either directly sequenced bidirectionally in 3730xl DNA Analyzer (Applied Biosystems) by Life Technologies, Shanghai, China, or cloned into pMD18-T vector (Takara) as previously described (Ge et al. 2012). Plasmids were extracted using the Miniprep Kit (Axygen) and then sequenced bidirectionally by Life Technologies.
All the DNA sequences obtained in this study had primer region sequences removed before analyses. DNA sequences or deduced amino acid sequences were blast against NCBI database. Multiple DNA sequences were aligned by the MegAlign program using the Clustal W method in Lasergene software suite (DNASTAR). Phylogenetic trees were generated using the neighbor-joining (NJ) algorithm implemented in Lasergene.
The representative nucleotide sequences of newly identified 16S rRNA and gltA genes of Anaplasma and rompB genes of SFG Rickettsiae in this study have been deposited in GenBank. The nucleotide sequence accession numbers were shown in the figures of phylogenetic analyses.
Results
Detection of Anaplasma in goats and sheep by nested PCR
To detect A. phagocytophilum, A. bovis, and A. centrale, the amplifications of partial 16S rRNA genes by nested PCRs were performed using species-specific primers. The prevalence of Anaplasma spp. in goats and sheep in the 10 farms is summarized in Table 1, which is illustrated in Figure 2A. Overall, 51% (51 of 100) of the tested samples were positive for Anaplasma, which distributed in eight farms, except A-1 and B-2. Fifteen percent, 49%, and 16% of the tested samples were positive for A. phagocytophilum, A. bovis, and A. centrale, respectively. E. chaffeensis was not detected in this study.
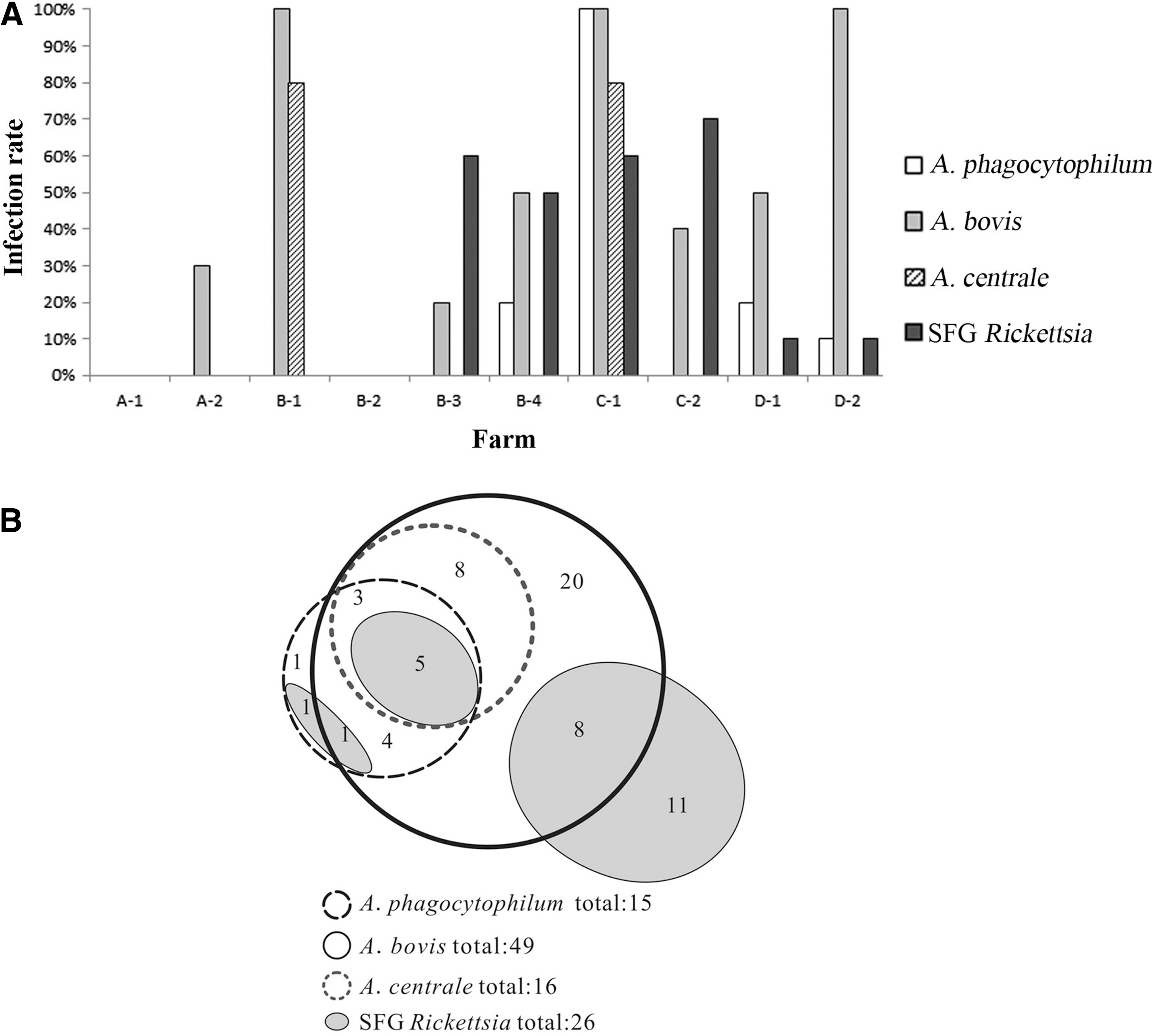
The infection of Anaplasma phagocytophilum, Anaplasma bovis, Anaplasma centrale, or spotted fever group (SFG) Rickettsia in tested samples from 10 farms.
Detection of SFG Rickettsiae in goats and sheep by nested PCR
To detect SFG Rickettsia infection in goats and sheep, the amplification of partial rickettsial rompB genes was performed by nested PCR. As shown in Table 1, 26% of tested samples were positive for SFG Rickettsiae, which were mainly from the four farms of B-3, B-4, C-1, and C-2 (Fig. 2A).
Coinfection
As shown in Figure 2B, eight of the tested samples were coinfected with A. phagocytophilum, A. bovis, and A. centrale. Thirteen samples were coinfected with A. phagocytophilum and A. bovis, 8 samples with A. phagocytophilum and A. centrale, and 16 samples with A. bovis and A. centrale. Fifteen samples were coinfected with Anaplasma and SFG Rickettsia. Five samples were coinfected with A. phagocytophilum, A. bovis, A. centrale, and SFG Rickettsia.
Sequence analyses of Anaplasma 16S rRNA genes
The detection of Anaplasma by PCR on the 16S rRNA genes was confirmed by the DNA sequencing of PCR amplicons. The 16S rRNA gene sequences of 15 A. phagocytophilum-positive samples were highly homologous to each other with identities from 99.5% to 100% (data not shown). The 16S rRNA gene sequences of 16 A. centrale-positive samples were 100% identical to each other (data not shown). The identities of 16S rRNA gene sequences for 49 A. bovis-positive samples vary from 98.8% to 100% (data not shown).
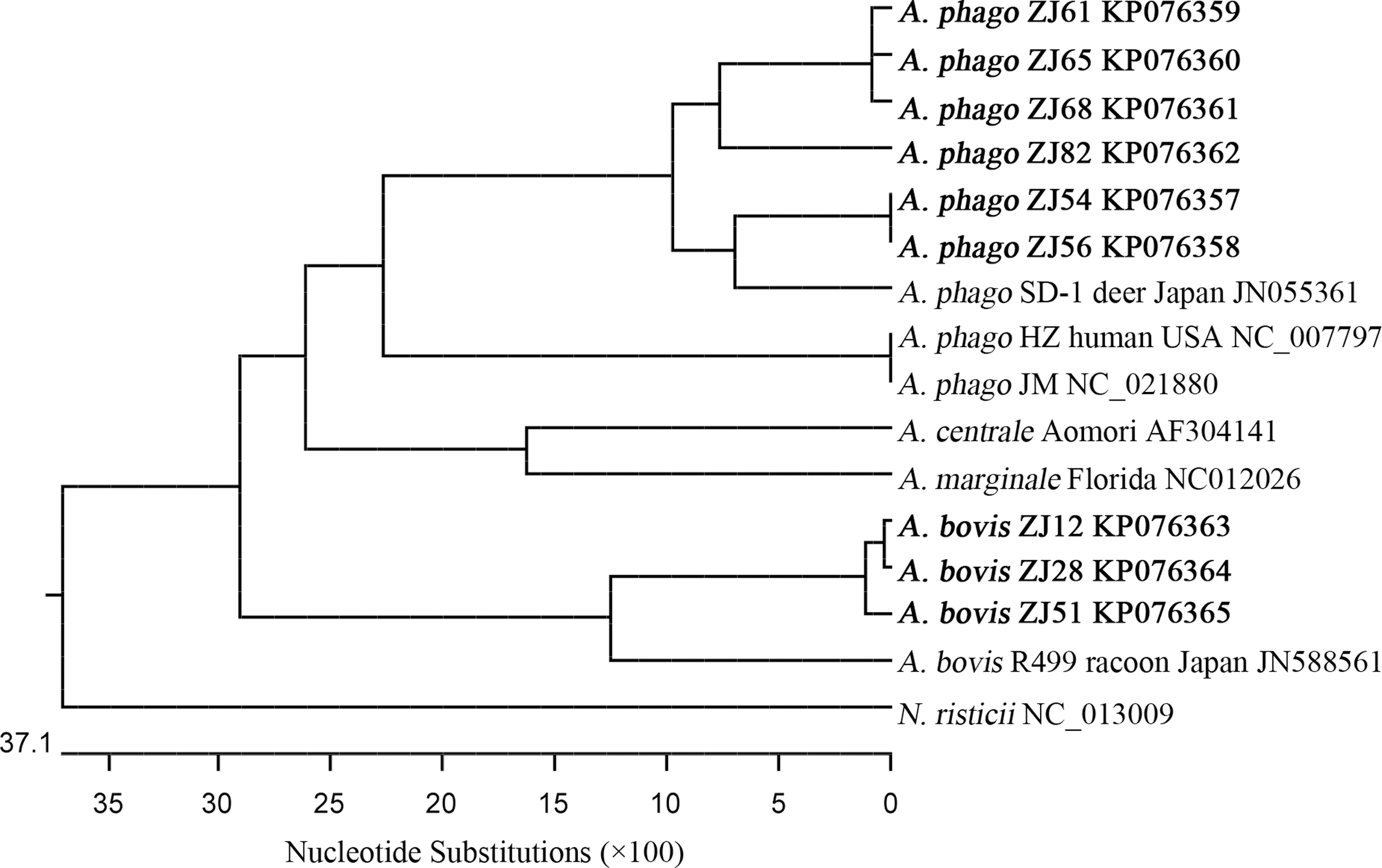
Longer fragments (>1300 bp) of Anaplasma 16S rRNA genes were amplified for selected 25 positive samples. The representative sequences have been deposited in GenBank and used to build the phylogenetic tree (Fig. 3). Blast results have shown that the Anaplasma 16S rRNA genes of ZJ56 and ZJ81 had 99% identity (with 100% query cover) with the sequences of strains from Japanese sika deer SD-1 (GenBank acc. no. JN055357) (Ybanez et al. 2012), Japanese sika deer SS33P-L (GenBank acc. no. AB196721) (Kawahara et al. 2006), Japanese cattle (GenBank acc. no. EU368728) (Ooshiro et al. 2008), and South Africa dog SA1076 (GenBank acc. no. AY570539) (Inokuma et al. 2005) and that the Anaplasma 16S rRNA genes of ZJ56 and ZJ81 had 98% identity (with 100% query cover) with the sequence of strain from Chinese Hubei goat YDH6 (GenBank acc. no. JN558811) (Liu et al. 2012). The Anaplasma 16S rRNA genes of ZJ56 and ZJ81, however, were not clustered with the strains from the Chinese Zhejiang rodent (GenBank acc. no. DQ458808) (Zhan et al. 2008), rodent from northeastern China (GenBank acc. no. DQ342324) (Cao et al. 2006), or human isolate A. phagocytophilum str. HZ USA (GenBank acc. no. NR_074113) (Dunning Hotopp et al. 2006).

Rooted neighbor-joining (NJ) phylogenetic tree of the Anaplasma 16S rRNA genes (>1300 bp) detected in this study and those published in GenBank.
The Anaplasma 16S rRNA genes of ZJ15 and ZJ98 had the highest 99% identities (with 100% query cover) with A. bovis Chinese Zhejiang goat Yu12 (GenBank acc. no. JN558829) (Liu et al. 2012). The Anaplasma 16S rRNA gene of ZJ66 had the highest 99% identities (with 100% query cover) with A. bovis Southwest China goat wlg-2 (GenBank acc. no. FJ389577) (Zhou et al. 2010), Japanese cattle (GenBank acc. no. EU368731) (Ooshiro et al. 2008), and Chinese Guizhou goat G49 (GenBank acc. no. JN558824, 1354 of 1359 bp) (Liu et al. 2012). The Anaplasma 16S rRNA gene of ZJ54 had 99% identity (with 100% query cover) with that from the Chinese Zhejiang goat Yu12 strain (GenBank acc. no. JN558829) (Liu et al. 2012). The Anaplasma 16S rRNA gene of ZJ69 had more variation and was mostly related (with 99% query cover) to Chinese Guizhou goat G1 (GenBank acc. no. JN558822) (Liu et al. 2012).
The Anaplasma 16S rRNA genes of ZJ23 and ZJ65 were clustered with the strain from southwestern China goat (GenBank acc. no. FJ389575) (with 100% query cover) (Zhou et al. 2010). The Anaplasma 16S rRNA genes of ZJ62, ZJ64, and ZJ68 had 99% identity (with 100% query cover) with those from A. centrale Japanese cattle Aomori strain (GenBank acc. no. AF283007) (Inokuma et al. 2001).
Sequence analyses of Anaplasma gltA genes
To look at the polymorphism of Anaplasma, the nucleotide sequences of partial Anaplasma gltA genes (413 bp) were obtained in nine representative positive samples. The sequence analyses of Anaplasma gltA genes are consistent with those of Anaplasma 16S rRNA genes in this study. As shown in Figure 4, the nine Anaplasma gltA gene sequences were clustered into two major clades. The Anaplasma gltA gene sequences of ZJ54 and ZJ56 are 100% identical (data not shown). The nucleotide and deduced amino acid sequences of ZJ54 and ZJ56 Anaplasma gltA genes had the highest identities of 88.3% (331/375) and 96% (119/124) with those of Japanese sika deer SD-1 strain (GenBank acc. nos. JN055361 and AER38795) (Ybanez et al. 2012), respectively. The nucleotide amino acid sequences of ZJ82, ZJ61, ZJ65, and ZJ68 Anaplasma gltA genes had the identities of 82.6% to 83.6% (373 bp in comparison) with those of Japanese sika deer SD-1 strain (Ybanez et al. 2012). The other cluster consists of the three samples of ZJ12, ZJ51, and ZJ28. By Blast search, the nucleotide sequences of Anaplasma gltA genes from ZJ12, ZJ51, and ZJ28 had no similar sequences, whereas their deduced amino acid sequences had the highest identities of 88% to 89% (124 amino acids in comparison) (GenBank acc. no. AEX07896) with those of A. bovis strain from the Japanese raccoon R499 (Sashika et al. 2011). The nucleotide sequences of Anaplasma gltA genes from ZJ12, ZJ51, and ZJ28 had identities of 78.8% to 79.6% (373 bp in comparison) with those from the same raccoon R499 strain (GenBank acc. no. JN588561) (Sashika et al. 2011).
Rooted NJ phylogenetic tree of the Anaplasma gltA genes detected in this study and those published in GenBank.
Sequence analyses of rickettsial rompB genes
The 26 rickettsial rompB sequences identified in this study fell into two major clusters by phylogenetic analysis (data not shown). The results of representative rickettsial rompB sequences (382 bp) are shown in Figure 5. One cluster included the 20 rickettsial sequences of samples such as ZJ50, ZJ60, ZJ66, ZJ72, ZJ76, and ZJ80. (data not shown). Blast results have shown that they have the highest 99% to 100% identities with the sequences from Rickettsia conorii str. Malish 7 (GenBank acc. nos. AE006914 and AF149110) (Ogata et al. 2000, Stenos and Walker 2000), South Korea human strain (GenBank acc. no. DQ003583) (Choi et al. 2005), and strain Indian tick typhus rickettsia (GenBank acc. no. AF123726) (Roux and Raoult 2000). In the other cluster were six rickettsial sequences of samples such as ZJ58, ZJ64, and ZJ99 (data not shown). The nucleotide sequences of the six samples have 99.5% to 100% identities to each other and their deduced amino acid sequences are 100% identical. Blast results have shown that the nucleotide sequences of the six samples have the highest identity of 99% to 100% with the sequences from Rickettsia rickettsii strains such as str. Iowa (GenBank acc. no. CP000766) (Ellison et al. 2008), str. Dv090589 (GenBank acc. no. GU395293) (Stromdahl et al. 2011), and str. R (GenBank acc. no. X16353) (Gilmore et al. 1989).
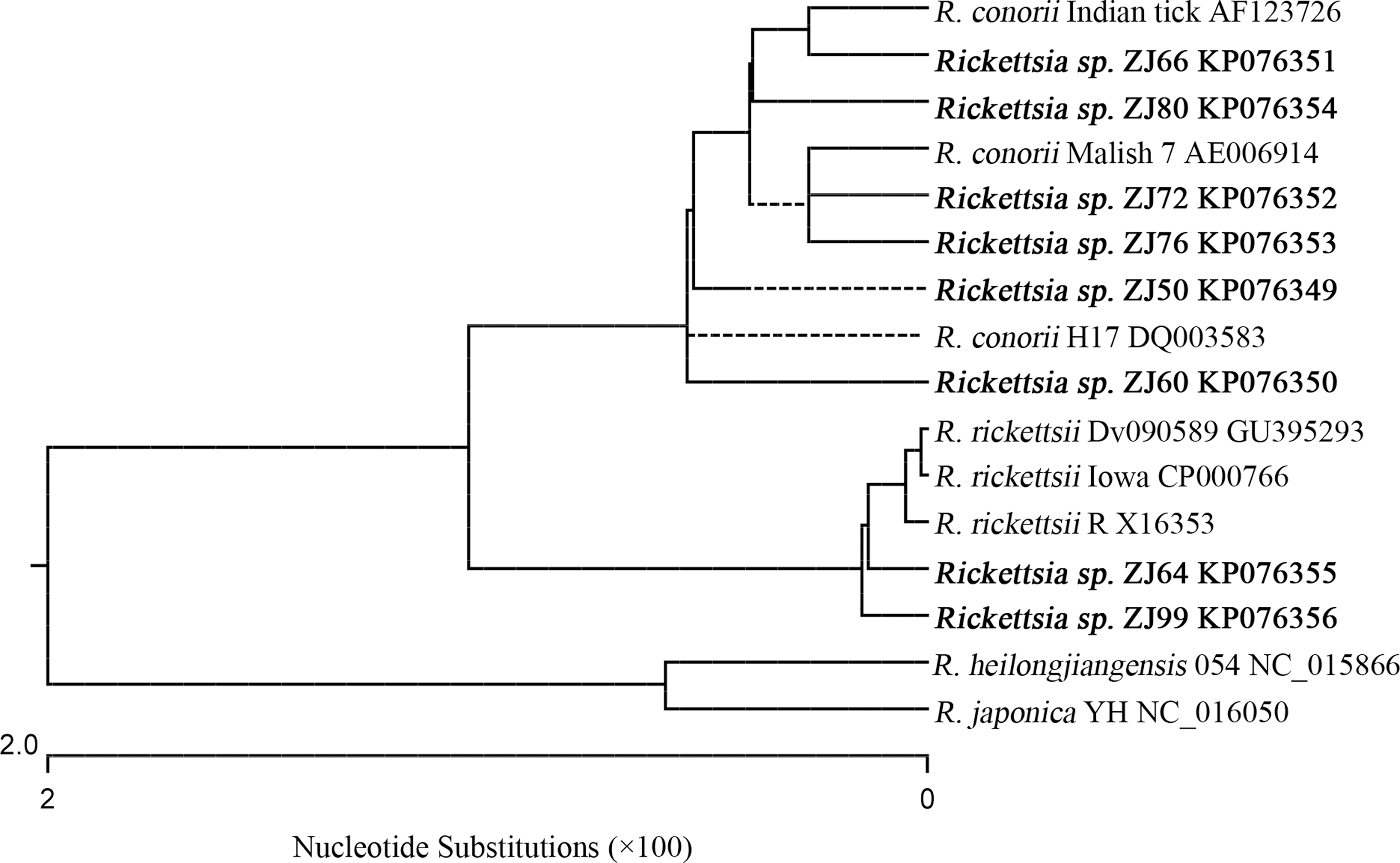
Rooted NJ phylogenetic tree of the SFG rickettsial rompB genes detected in this study and those published in GenBank.
Discussion
The present study detected the presence of Anaplasma in goats and sheep in Zhejiang province in China, which includes the three species of A. phagocytophilum, A. bovis, and A. centrale. Among these Anaplasma species, the infection rate of A. bovis (51%) was the highest, suggesting that A. bovis was the dominant Anaplasma species prevalent in goats and sheep in southeastern China. The infection rate for A. bovis determined in this study is close to that (49.6%) in goats from central and southern China in Liu's study (Liu et al. 2012). Compared with the 16S rRNA sequences of A. phagocytophilum and A. centrale detected in this study, those of A. bovis are more diverse. So far, except the 16S rRNA gene, little genetic information on A. bovis is available. The A. bovis gltA sequence data obtained in this study contribute to the molecular studies on A. bovis.
The A. phagocytophilum detected in this study was subclustered to the Japanese SD-1 strain from sika deer (Ybanez et al. 2012), which was relatively distant to A. phagocytophilum from rodents detected previously in the same province, Zhejiang (Zhan et al. 2008) or northeastern China (Cao et al. 2006). These results suggested that more than one sequence of A. phagocytophilum was present in Zhejiang and that no geographic segregation was observed. The infection rate of A. phagocytophilum in goats and sheep in this study was 15%, which was close to that (14.5%) in goats from central and southern China in Liu's study (Liu et al. 2012).
The A. centrale detected in this study has the highest identity to that reported previously in Japan (Inokuma et al. 2001). The Anaplasma 16S rRNA genes of ZJ23 and 65 have less than 99% identity with published sequences. These Anaplasma variants were most closely related to A. centrale. More genetic characteristics of these Anaplasma variants wait for future study.
This study identified the presence of SFG Rickettsia in domestic animals in southeastern China. The SFG Rickettsia spp. detected in this study potentially includes the two species, i.e., R. conorii and R. rickettsii. R. rickettsii was first found in North America. It has been molecularly or serologically reported in several Asian countries, e.g., Japan, Korea, and Thailand (Uchida et al. 1985, Suksawat et al. 2001, Kim et al. 2006). Although the rickettsial rompB gene sequence is not sufficient to define rickettsial species, it has been shown to be able to discriminate different Rickettsia spp., including R. conorii in a clinical or field study (Choi et al. 2005, Palomar et al. 2012). Zhan et al. (2009) have previously detected SFG Rickettsia in rodents in Zhejiang by PCR, but no sequencing data were available. Further study on the characteristics of SFG Rickettsiae detected in this study is desired.
All the samples previously detected positive for the tick-borne microbe, Theileria (Ge et al. 2012), were also positive for Anaplasma in this study, which reflects a coinfection of organisms from these two genera. The detection of Anaplasma and Rickettsia in this study further alerts people about the potential threat of tick-borne pathogens to the health of livestock as well as humans in the study areas. The coinfection of tick-borne pathogens may facilitate the lateral gene transfer between organisms and contribute to the occurrence of variation, which deserves study in the future. Samples from farms A-1 and B-2 were negative for all the tested pathogens, suggesting of no or few infectious tick vectors transmitting the pathogens in the study areas. It would be very interesting to study the involvement of vectors transmitting the Rickettsiales as well as Theileria in these areas in the future. More knowledge on the genetic characteristics of these organisms, pathogenicity, and involved hosts is also expected.
Footnotes
Acknowledgments
The authors thank Dr. Bohai Wen from Beijing Institute of Microbiology and Epidemiology, China, for providing the R. heilongjiangensis 054 strain DNA control. This work is supported by the project from the Scientific Research Foundation for the Returned Overseas Chinese Scholars, State Education Ministry, China; the Pujiang Talent program 09PJ1410100 from Science and Technology Commission of Shanghai Municipality, China; the project SKLVEB2010KFKT012 from the State Key Laboratory of Veterinary Etiological Biology, Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences.
Author Disclosure Statement
No competing financial interests exist.