
Abstract
Staphylococcus aureus in food is a consequence of inadequate hygienic handling and processing, posing a potential risk to public health. The current study aimed to characterize virulence factors, as well as antimicrobial resistance of Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) isolated from retail chicken products and hand swabs from vendors in Egypt. In addition, genetic relatedness of the isolates from chicken and humans was evaluated by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) using protein A as a target. A total of 110 samples were collected from chicken products (n = 80) and vendors (n = 30). Overall, 30 (37.5%) chicken products samples were positive for S. aureus, whereas hand swabs from meat handlers revealed that 18 (60%) were positive. Ten MRSA strains were characterized by the presence of the mecA gene, comprising seven isolates from chicken and three from humans. Virulence-associated factors were evaluated by PCR, revealing that 31.3% of S. aureus isolates harbored the Panton–Valentine leukocidin (PVL) gene, whereas 10.4% were positive for the sea and sed genes each, and only two isolates were positive for γ-hemolysin–associated gene. Genotyping using spa PCR-RFLP showed identical restriction banding patterns of MRSA isolates of human and chicken meat origin, indicating the genetic relatedness of the isolates. To the best of our knowledge, this is the first study to characterize PVL-positive MRSA from chicken products and to utilize spa-RFLP for evaluating the genetic relatedness between MRSA of human and chicken origin in Egypt.
Introduction
T
Resistance to antibiotics is an increasing concern worldwide and is associated mainly with uncontrolled usage of antimicrobials for treatment or as growth promoters in food animals (Barber et al. 2003). S. aureus resistant to penicillinase-resistant pencillins is referred to as methicillin-resistant S. aureus (MRSA). Resistance of methicillin is mediated by the mecA gene, which encodes penicillin-binding protein 2a, with a decreased binding affinity for β-lactams (Chambers 1997). Because of its multidrug resistance pattern, MRSA is considered a public health concern, especially for immunocompromised patients (Pesavento et al. 2007, Kraushaar and Fetsch 2014). Panton–Valentine leukocidin (PVL) is one of the important exotoxins causing severe skin and soft tissue infections and necrotizing pneumonia (Kraushaar and Fetsch 2014). The association of PVL with progressive, hemorrhagic, and necrotizing pneumonia has been reported in cases of community-acquired infection with MRSA (Gillet et al. 2002). γ-Hemolysin is also a toxin causing damage to erythrocytes (Prevost et al. 1995). The gene is encoded by the hlg locus, and previous reports have described the gene in more than 99% of S. aureus strains (Prevost et al. 1995). Production of enterotoxins by S. aureus is another virulence-associated factor that has an impact on the pathogenicity of the isolates. Eleven antigenic types of enterotoxins (staphylococcal enterotoxin type A [SEA] to SE type E [SEE]) are recognized; of them, SEA is considered the most commonly involved enterotoxin with human infection (Balaban and Rasooly 2000).
Typing of MRSA is essential for tracing the source of infection; therefore, typing methods should have the advantages of typeability, reproducibility, and good discriminatory power (Wichelhaus et al. 2001). Different molecular methods, e.g., pulsed-field gel electrophoresis (PFGE), have been described as the gold standard typing method; however, PFGE is not widely used due to expense, time required, and sophisticated application (Wichelhaus et al. 2001). Molecular typing of S. aureus using polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) has been shown to be of sufficiently high accuracy and corresponds to PFGE typing (Omar et al. 2014).
This study aimed to assess retail chicken products sold in Egypt as a potential vehicle for S. aureus and MRSA. Antimicrobial resistance profiling and molecular characterization of some virulence factors of the isolates obtained were carried out. Moreover, the genetic relatedness of MRSA isolates of human and chicken origin was determined by protein A (spa) PCR-RFLP.
Materials and Methods
Sampling and isolation of S. aureus
A total of 80 chicken products' samples (40 raw chicken breast fillet, 20 sliced luncheon meat, and 20 chicken nuggets) were collected randomly from local pluck-shop markets and outlets at Sharkia Governorate, Egypt. Moreover, 30 hand swabs from chicken handlers at the pluck-shop markets and outlets were collected. Twenty-five grams from each sample were homogenized with 225 mL of buffered peptone water (BPW; cat. no. CM509, Oxoid, Adelaide, Australia) for pre-enrichment and incubated at 37°C for 24 h. A loopful from the pre-enriched culture homogenate in BPW was inoculated onto Baird–Parker agar medium (cat. no. CM0275, Oxoid, Adelaide, Australia) and incubated at 37°C for 24 h (ISO-6888-1, 1999). In addition, serial dilutions were prepared for S. aureus count. One suspected S. aureus colony from each plate was subjected to Gram staining and biochemical identification (catalase, coagulase, oxidase, and DNase). S. aureus identification was then confirmed by amplification of the species-specific nuc gene (Gao et al. 2011).
Antimicrobial susceptibility test
A disc diffusion test was carried out to detect the minimum inhibitory concentration (MIC) of S. aureus isolates against 14 antimicrobials according to National Committee for Clinical Laboratory Standards (NCCLS 2013). The antibiotics that were used to evaluate resistance are listed in Table 2 (below).
Molecular characterization and spa-RFLP
S. aureus isolates were subjected to PCR for the detection of mecA (Zhang et al. 2004), PVL (Lina et al. 1999), γ-hemolysin (Kumar et al. 2009), and enterotoxin encoding genes (sea, sec, and sed) (Mehrotra et al. 2000). Moreover, MRSA strains isolated from different sources were subjected to spa-PCR-RFLP for genotyping (Omar et al. 2014). Extraction of the bacterial DNA was carried out using a QIAamp DNA Mini Kit according to the manufacturer's guidelines (Qiagen, Germany, GmbH). For restriction, HindIII enzyme was used according to the protocol described by Omar et al. (2014). The synthesized primers and the amplicon sizes are summarized in the Table S1 (Supplementary Data are available at
Results
Biochemical identification revealed that 67 isolates were suspected S. aureus (data not shown). Only biochemically suspected S. aureus isolates confirmed by nuc gene PCR were included in the study (n = 48). A total of 110 samples comprising 40 chicken breast fillet, 20 sliced luncheon meat, 20 chicken nuggets, and 30 hand swabs from vendors were examined. Overall, 30 (37.5%) chicken samples were positive for S. aureus, with isolation rates of 42.5%, 40%, and 25% from chicken fillet, nuggets, and sliced luncheon meat, respectively (Table 1). The difference in the contamination rate of the investigated products with S. aureus was insignificant (χ1 2 = 1.8, p = 0.4). Examination of hand swabs collected from vendors revealed that 18 (60%) were positive for S. aureus (Table 1). The total S. aureus counts in chicken products were 3.4 × 104 ± 8.4 × 103 (chicken fillet), 1.1 × 104 ± 3.3 × 103 (nuggets), and 6.3 × 104 ± 1.5 × 104 (sliced luncheon meat) (Table 1). Out of the examined isolates from chicken (n = 80) and vendors (n = 30), the mecA gene specific for MRSA was detected in seven (8.8%) and three (10%) samples, respectively (Table 1).
Number (proportion, 95% confidence interval).
MRSA, methicillin-resistant S. aureus; CFU, colony-forming units; SE, standard error.
The results of the antibiotic resistance of 48 S. aureus isolates of chicken and human origin showed that all of the isolates (100%) were sensitive to vancomycin, and only 7.8% resistance was observed against ofloxacin (Table 2). High resistance was detected against doxycycline (64.6%) and amoxicillin (60.4%). However, resistance rates ranging from 18.8% and 47.9% were reported to the other examined antimicrobials (Table 2).
MRSA, methicillin-resistant S. aureus.
Acquisition of virulence factor–associated genes was investigated by molecular amplification of PVL-, γ-hemolysin–, and enterotoxin-associated genes (sea, sec, and sed) Fig. S1. The results showed PVL, γ-hemolysin–, sea-, and sed-associated genes were detected, respectively, in 31.3%, 4.2%, 10.4%, and 10.4% of the S. aureus isolates obtained (Table 3). The comparable occurrence of PVL, γ-hemolysin–, sea-, and sed-associated genes in MRSA versus non-MRSA isolates was 90% vs. 15.8%, 0 vs. 5.3%, 40% vs. 2.6%, and 30% vs. 5.3%, respectively. None of the examined S. aureus isolates (MRSA or non-MRSA) was positive for the sec-associated gene (Table 3).
MRSA, methicillin-resistant S. aureus.
Genotyping of the spa region in 10 MRSA isolates (seven of chicken and three of human origin) showed that 80% of the identified MRSA isolates produced a single band of 1200 bp (type S1), whereas only 20% produced no bands (Fig. 1). The absence of PCR product is considered a separate type, namely S2. Restriction of the produced amplicon by the HindIII enzyme produced two banding patterns of 240 bp and 696 bp (Fig. 1). The typeability of the method was calculated to be 80%.
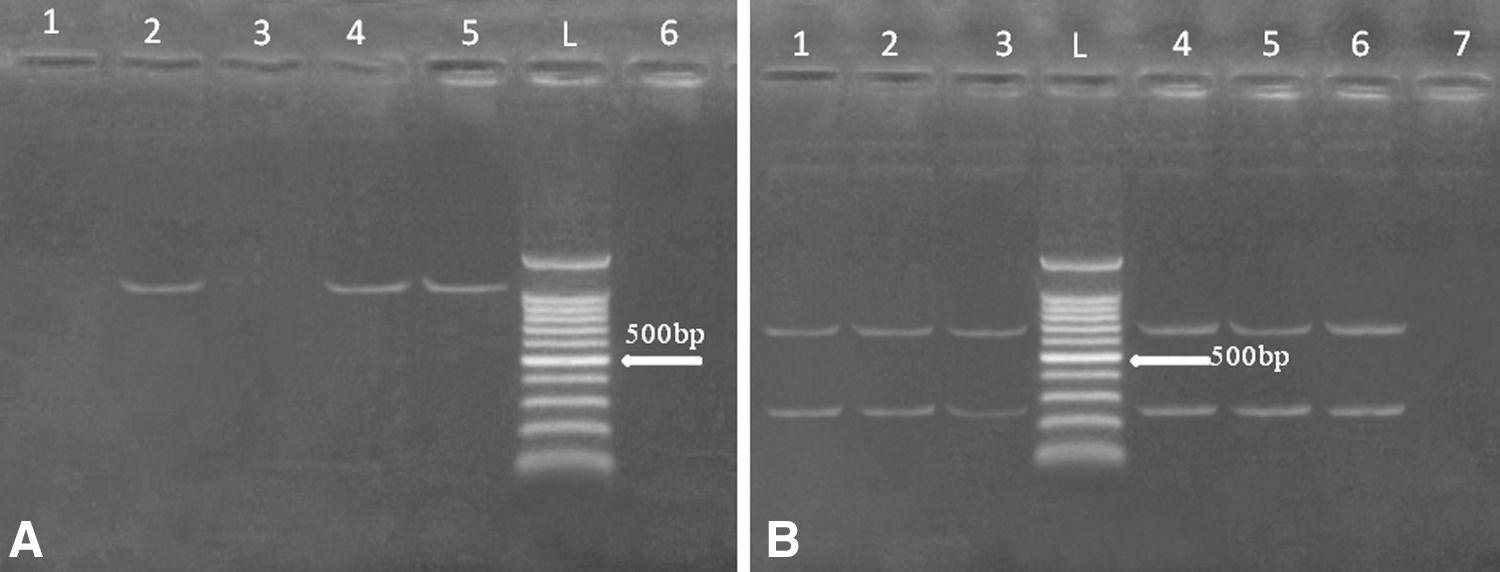
Exemplar for 1% agarose gel of spa-RFLP. Lane L, 100-bp ladder. (
Discussion
Contaminated chicken meat is considered an important source of foodborne pathogens posing risk for zoonotic transmission (Osman et al. 2015). In the current study, the proportion positive for S. aureus in chicken products' samples was 37.5% (Table 1). In Egypt, S. aureus isolation rates of 6% (EI-Jakee et al. 2013, Osman et al. 2015) and 77.6% (Karmi 2013) from chicken meat samples have been reported. The difference between the isolation rate obtained and those previously reported in Egypt could be attributed to the identification of S. aureus isolates using only biochemical identification in the aforementioned three studies. Phenotypic identification of staphylococci has been reported to be unsatisfactory, irreproducible, and unreliable (Heikens et al. 2005). Moreover, the variation could be attributed to the sample size, location, and hygienic measures posed by chicken meat handlers (Bhargava et al. 2011). Comparable isolation rates were reported in Japan (34.2%; Ogata et al. 2012), the United States (42.1%; Abdalrahman et al. 2015), and South Africa (44%; Oguttu et al. 2014).
As illustrated in Table 1, S. aureus contamination was found to be higher in chicken breast fillet (42.5%) and nuggets (40%) than in sliced luncheon meat (25%); however, the difference between the three products was insignificant. S. aureus was recorded in 46.2% of chicken fillets in Croatia (Kozačinski et al. 2006), 75% of nuggets in Italy (Pepe et al. 2006), and 40% of luncheon meat samples in Egypt (Karmi 2013). The lower contamination rate of sliced luncheon meat than chicken fillet and nuggets might be attributed to the complete heat treatment of luncheon meat during processing. Although there was a low contamination rate of sliced luncheon meat, the potential of handling after cooking is suspected, which, in turn, poses a risk for human infection (Oguttu et al. 2014).
MRSA is a potential cause of hospital-acquired infection; however, community-acquired infections are also of growing concern (Tiemersma et al. 2004, Witte et al. 2004). The isolation rate of MRSA from chicken products in the present study was 8.8% (Table 1). Similarly, 7.1% of chicken meat samples were positive for MRSA in Korea (Cho et al. 2014). Variable isolation rates of MRSA from chicken meat have been reported in different studies (de Boer et al. 2009, Fessler et al. 2011, Hanson et al. 2011, Agerso et al. 2012, Chairat et al. 2015). The results obtained revealed that contamination of poultry meat during slaughter is assumed to be the most important source for infection of human consumers. Such an assumption was supported by the reported observation in The Netherlands that 8% of poultry were MRSA positive before slaughtering; however, after slaughtering, 35% of poultry meat was contaminated (Mulders et al. 2010).
Infected food handlers are responsible for foodborne illnesses due to bacterial pathogens (Centers for Disease Control and Prevention 2000). It has been estimated that more than 50% of healthy individuals have a high prevalence of S. aureus on the hands and fingertips and that food handlers have been involved in outbreaks (Jones et al. 2002, Le Loir et al. 2003, Lues et al. 2006). The results showed that 60% of hand swabs collected from food handlers at retail markets were positive for S. aureus, with MRSA comprising 10% (Table 1). S. aureus was isolated from 57.5% of food handlers in Botswana (Loeto et al. 2007) and 38.3% in Nigeria (Okareh and Erhahon 2015). Moreover, in Nigeria, 48.8% of hand swabs from meat handlers were positive for MRSA, and the authors attributed such a high isolation rate to the observed unhygienic handling of meat (Nnachi et al. 2014). On the contrary, MRSA was not recovered from hand swabs of meat handlers in The Netherlands as a consequence of the continuous active surveillance against antibiotic-resistant microorganisms (de Jonge et al. 2010).
According to the legal requirements of the Egyptian Organization for Standardization and Quality Control (EOSQC 2005), the permissible level of S. aureus in poultry meat varies from one product to another. In chicken fillets, S. aureus should be less than 102 colony-forming units (CFU)/gram, whereas partially and completely heat-treated chicken products (nuggets and luncheon meat, respectively) should be S. aureus free. The results obtained showed that the mean S. aureus count in chicken breast fillet, nuggets, and sliced luncheon meat are beyond the permissible limits of EOSQC (Table 1), thus indicating that chicken products in retail at the study area pose risk to human consumers. Lower S. aureus counts of 2.74 ± 0.56 log CFU/gram were reported from breast fillet in Croatia (Kozačinski et al. 2006) and >103 in ready-to-eat chicken meat in South Africa (Oguttu et al. 2014), while, in The Netherlands, colony counts in chicken meat products were found below 10 CFU/gram (de Boer et al. 2009).
The emergence of antimicrobial-resistant S. aureus has been associated with the uncontrolled use of drugs such as growth promoters in animals; hence, there should be a restriction of usage at the farm level (Huber et al. 2010, Buyukcangaz et al. 2013). In the present study, 64.6% and 60.4% of the isolates were resistant to doxycycline and amoxicillin (Table 2). In accordance with these results, S. aureus isolates have shown increased resistance to tetracyclines and penicillins in different studies (Aydin et al. 2011, Pu et al. 2011, Buyukcangaz et al. 2013). The spread of penicillin resistance has been found to be rapid due to plasmid transfer and is most reported among isolates of food origin (Aydin et al. 2011, Buyukcangaz et al. 2013). S. aureus isolates of human origin were significantly more resistant to cefoperazone than isolates of chicken origin (χ1 1 = 5.34, p = 0.02). The higher resistance of human isolates could be attributed to the common use of cefoperazone in treatment of dairy cattle infected with mastitis, which consequently results in more exposure of human consumers to resistant S. aureus from dairy sources (Fessler et al. 2014). Interestingly, all S. aureus isolates in the present study were sensitive to vancomycin, thus concurring with previously reported results (Buyukcangaz et al. 2013, Cho et al. 2014, Kim et al. 2015).
Characterization of some virulence-associated genes was essential to investigate the potential of the isolates to pose risk to human consumers. Of 48 S. aureus isolates, the PVL-associated gene was found in 31.3%, whereas 90% of MRSA isolates were PVL positive (Table 3). In agreement with the current study, all MRSA isolates from retail meat were PVL positive in the United States (Pu et al. 2009, Bhargava et al. 2011), whereas 37.2% of S. aureus isolates from clinical cases were positive for PVL (Lina et al. 1999). In Egypt, 19.04% PVL positive MRSA isolates were characterized from patients at Zagazig City, Egypt (Enany et al. 2010). Other studies reported that MRSA isolates from retail meat were negative for the PVL-associated gene (Fessler et al. 2011, Hanson et al. 2011, Ogata et al. 2012). The significance of PVL-positive MRSA isolates was associated with their ability to produce skin infections and pneumonia (Liassine et al. 2004, Labandeira-Rey et al. 2007).
Although γ-hemolysin previously was reported to be produced by more than 99% of S. aureus (Prevost et al. 1995), only two isolates were positive in the present study (Table 3). Likewise, Li et al. (2015) reported that none of the S. aureus isolates from retail foods in China were positive for the hlg gene. Contrary to our results, γ-hemolysin–associated gene was detected in all MRSA isolates from human cases (Lina et al. 1999).
S. aureus isolates of poultry origin mainly produce toxins C and D; however, production of toxins A and B was reported recently in isolates from raw chicken meat (Kitai et al. 2005). In the present study, 10.4% of S. aureus isolates were positive for both sea- and sed-associated genes (Table 3). In Japan, Kitai et al. (2005) reported that 78 S. aureus isolates were enterotoxigenic, of which, 17.9% and 2.6% were found to produce SEA and SED, respectively. Enterotoxins were reported also in 29.5% of S. aureus isolated from meat and meat products in China (Jiang et al. 2001).
In the present study, 40% and 30% of MRSA isolates were found to harbor sea and sed genes, respectively (Table 3). Similarly, Corrente et al. (2005) reported that 71 MRSA isolates of human origin in Italy were able to synthesize SEA, whereas, in contrast, SED was the most frequently synthesized toxin among MRSA isolates from foods of animal origin in Italy (Normanno et al. 2007). Most of MRSA isolates (60%) were shown to harbor more than one of the investigated virulence factors, whereas only two non-MRSA isolates showed the simultaneous presence of two virulence determinants. These results indicate that MRSA isolates pose more risk to human consumers compared to non-MRSA isolates (Kim et al. 2015).
The spa gene encoding for protein A was chosen for typing the isolates in the current study on the basis of the previously reported heterogeneous nature indicating variability among the examined strains (Wichelhaus et al. 2001). The results obtained showed that a single band of 1200 bp was produced by 80% of MRSA isolates, whereas 20% of the strains had no bands (Fig. 1). These results were consistent with Omar et al. (2014), who reported that PCR-RFLP of the spa gene produced a single band (type S1) in 84% of MRSA isolates; 5.2% were of type S4 that had no PCR products. Moreover, the authors reported the same banding patterns after restriction and 92% typeability of the method. The absence of the spa gene was also reported in 3.8% (Shakeri et al. 2010) and 5% (Adesida et al. 2006) of S. aureus isolates. These observations were supported by the speculation that the spa gene is not consistently expressed in all S. aureus isolates (Frénay et al. 1994).
The isolation of S. aureus with long spa bands (1200–1392 bp) has been reported to have a role in adherence to respiratory epithelium (Omar et al. 2014). The current study reports that spa bands from MRSA isolates of human and chicken meat origin are long enough (1200 bp) to adhere to respiratory epithelium in case of infection. In accordance, different studies have also reported the amplification of the spa gene in S. aureus producing long amplicons (Mehndiratta et al. 2009, Shakeri et al. 2010, Soltan Dallal et al. 2010). Two bands of 240 bp and 696 bp were produced by all MRSA isolates after restriction (Fig. 1). The identical restriction banding pattern of MRSA isolates of human and chicken meat origin indicated the genetic relatedness of the isolates. Interestingly, the current results showed that the seven MRSA isolates were from the shops where MRSA-positive vendors were identified. Such findings highlighted the possibility of MRSA transmission between meat handlers and chicken meat. The potential of contaminated meat consumption to produce risk for S. aureus infection to consumers was suggested by Ogata et al. (2012), who identified identical genetic fingerprint between S. aureus isolates of human and meat. Also, MRSA isolates of chicken origin were closely related to animal and human isolates in Korea (Lee 2003).
Conclusion
Although there was a low isolation rate of MRSA from the chicken products examined, this rate could present a risk to consumers. To minimize the spread of MRSA by contaminated raw foods, handlers should follow appropriate hygienic measures during the handling and preparation of food. Moreover, knowledge about the sources, mode of transmission, and prevention tools is required. To the best of our knowledge, this is the first study to characterize PVL-positive MRSA from chicken products and to use spa-RFLP for evaluating the genetic relatedness between MRSA of human and chicken origin in Egypt.
Footnotes
Acknowledgment
The authors express sincere gratitude to Rasha Mahmoud Hebesh, a Master student at the Food Control Department, Faculty of Veterinary Medicine, Zagazig University, for her valuable support during the bacteriological examination part of the current work.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.