
Abstract
Herpesviruses (HVs) can cause asymptomatic, benign, or fatal infections in a variety of animal species. However, the prevalence and phylogenetic characteristics of HVs in rodents and shrews in China are poorly understood. We thus performed a molecular detection and phylogenetic analysis of rat and shrew HVs in southern China between 2012 and 2014. Seventeen (6.7%) of 255 rectal swab specimens from rats and six (6.7%) of 90 rectal swab specimens from shrews tested positive for HVs. Phylogenetic analysis revealed that rodent and shrew HVs detected in this study were species specific, clustering in the Betaherpesvirinae and Gammaherpesvirinae clade. Novel Macavirus was detected in Rattus norvegicus (RN/13YX52/24 and RN/14HC50) and gammaherpesviruses in Suncus murinus (SM/14BY7/16/20/97/99/106).These findings have contributed to our understanding of the taxonomy, phylogeny, and biology of HVs.
Introduction
T
The order Rodentia is the largest group of mammalian species, accounting for 40% of all mammal species (Huchon et al. 2002). There are 2200 living rodent species, including mice, rats, voles, prairie dogs, beavers, and guinea pigs, known to act as vectors for more than 60 known human infectious diseases (Meerburg et al. 2009). Some species of rodents are commensal, living in close association with humans and offering numerous opportunities for the transmission of infectious disease pathogens through their urine or feces or through their arthropod ectoparasites, such as ticks, mites, and fleas. In southern China, there are three predominant commensal rodent species, including Rattus norvegicus, Rattus flavipectas, and Mus musculus. Rattus lossea Swinhoe, the most widely distributed field rodent species in southern China, has also been found to live in close association with humans in suburban locations.
Shrews are small insectivorous mammals that are commonly found in southern China. In the surveillance program of rodent-borne pathogens in southern China, we have collected a considerable number of shrews, which have been identified as the Asian house shrew (Suncus murinus), commonly found near human residences. Similar to rodents, some shrew species are recognized as reservoir hosts of some zoonotic diseases (Hsieh et al. 2010, Yashina et al. 2010, Sasaki et al. 2014).
Investigation of the HVs in rodents and shrews and their epidemiological and phylogenetic characteristics would significantly advance our understanding of the animal host ranges of HVs and the zoonotic potential of these animals. In an earlier study by Bruggeman et al. (1982), the cytomegalovirus-like particles were identified in wild rats by using light and electron microscopy. Recently, four studies (Ehlers et al. 2007, Teterina et al. 2009, Hughes et al. 2010, Ettinger et al. 2012) have reported novel rodent HV sequences in Europe, which mainly belonged to the Betaherpesvirinae and Gammaherpesvirinae subfamilies. However, rodent-derived HVs have not been reported in mainland China.
With interests in investigating the viruses that spread from animals to humans, we specifically targeted rodents and shrews that had a long history of living in close proximity to human populations and report the prevalence and phylogenetic characteristics of HVs detected from rodents and shrews in four geographic regions in Guangdong and Fujian provinces, southern China.
Materials and Methods
Ethics statement
The study protocol has been reviewed and approved by the Animal Ethics and Welfare Committee of the School of Public Health and Tropical Medicine, Southern Medical University. All animals were treated in strict accordance with the guidelines for the Laboratory Animal Use and Care from Southern Medical University and the Rules for the Implementation of Laboratory Animal Medicine (1998) from the Ministry of Health, China. All surgical procedures were performed under ether anesthesia in efforts to minimize suffering.
Sample collection
Rodents and shrews were captured around houses and markets using well-designed iron cage traps (280 × 120 ×100 mm) (Yue-zong Co Ltd.) with fried baits (fried breadstick and peanuts) in Tong-an (latitude: 24.73°N; longitude: 118.15°W) and Haicang (latitude: 24.47°N; longitude: 117.98°W) districts of Xiamen city and Yuexiu (latitude: 23.13°N; longitude: 113.27°W) and Baiyun (latitude: 23.17°N; longitude: 113.27°W) districts of Guangzhou city in southern China, between 2012 and 2014. The captured animals were euthanized with diethyl ether to collect rectal swabs. Rectal swabs were placed in 1000 μL of viral transport medium (Earle's balanced salt solution consisting of 0.2% sodium bicarbonate, 0.5% bovine serum albumin, 200 μg of vancomycin/L, and 160 U of nystatin/L). The samples then were transported to the laboratory and stored at −80°C until use.
Identification of rodent and shrew species
Rodent (Schlegel et al. 2012) and shrew (Kang et al. 2011) species identification was confirmed by morphological identification combined with amplification and sequencing of the cytochrome B (cytB) gene, which is a commonly applied technique in archeology (Linacre and Lee 2005). The cytB sequences of rodent and shrew detected in our study were compared with reported sequences in the GenBank database using the Basic Local Alignment Search Tool (BLAST). The taxonomy results of each rodent and shrew were determined based on the most similar hit searched by BLAST.
DNA extraction and PCR screening for HVs
Viral DNA from 200 μL of sample in the transport medium was extracted using stool DNA extraction kits (OMEGA) according to the manufacturer's instructions. Purified DNA was eluted in 50 μL of elution buffer. For initial detection of HVs, a nested PCR amplification targeting the highly conserved amino acid motifs in the DNA-directed DNA polymerase (DPOL) gene of HVs was performed by using the consensus primer sets (ILK, DFA, TGV, KG1, and IYG), according to the consensus-degenerate hybrid oligonucleotide primers program (van Devanter et al. 1996). Second-round products were found to be 179–232 bp in length.
For amplification of the diverse glycoprotein B (gB) sequences of Betaherpesvirinae, three different degenerate/deoxyinosine-substituted (deg/dI) nested primer sets were used—namely CM-gB1, CM-gB2, or CM-gB3 (Ehlers et al. 2007). Second-round amplification products were ∼225, 265, and 280 bp in length, respectively. For amplification of Gammaherpesvirinae, the deg/dI nested primer set, RH-gB, was used (Ehlers et al. 2007), targeting at the gB genes. Second-round amplification products had a calculated length of ∼467–516 bp.
Two microliters of DNA were added to form a 25 μL PCR system using the GoTaq Green Master Mix (Promega) according to the manufacturer's protocol. The procedures were identical to those described previously (Ehlers et al. 2007). The positive sample (human herpesvirus 6) and negative control (sterile water) were included in each reaction. A 5-μL sample of the final PCR product was separated by electrophoresis on 1% agarose gels stained with ethidium bromide.
Sequence and phylogenetic analysis
Sequence editing and identity calculations were conducted using BioEdit, version 7.0.4 (
Nucleotide sequence accession numbers
The nucleic acid sequences of herpesvirus DPOL, betaherpesvirus gB, and gammaherpesvirus gB genes reported in this study were deposited in the GenBank database under accession numbers, KT591375-KT591396, KT591352-KT591354, and KT591355-KT591374, respectively. For genetic analysis, other HV genomes were retrieved from the GenBank, including Alcelaphine herpesvirus 1 (AlHV-1, NC_002531), Ateline herpesvirus 3 (AtHV-3, NC_001987), Bovine herpesvirus (BoHV-4, NC_002665; BoHV-6, NC_024303; BLHV, AF327830), Bandicota indica rhadinovirus 4 (BIRV, EF128043.1), Canine herpesvirus (CaHV-1, AY949827), Equid herpesvirus 2 (EHV-2, HQ247792), Eptesicus serotinus rhadinovirus 1 (ESRV-1, DQ788623), feline herpesvirus 1 (FeHV-1; AJ224971), Human herpesvirus (HHV-1, X14112; HHV-2, Z86099; HHV-4, LN827580; HHV-5, M17209; HHV-6, M63804; HHV-7, NC_001716; HHV-8 GQ994935), Macacine herpesvirus 4 (MaHV-4, NC_006146), Miniopterus schreibersii herpesvirus/11HN96 (MS/11HN96, KR261847), Murid herpesvirus (MuHV-2, AY728086; MuHV-8, KP202868; MuHV-4, NC_001826), Mustelid herpesvirus 1 (MustHV-1, AF376034.1), Myotis ricketti herpesvirus (MyHV-1, JN692429; MyHV-2, JN692430), Ovine herpesvirus 2 (OvHV-2, NC_007646), Panine herpesvirus 1 (PaHV-1, AF306944), Rhinolophus blythi herpesvirus/13YF104 (RB/13YF104, KR261853), and Rattus exulans cytomegalovirus 1 (RECV-1, EF125071).
Results
Detection of HVs in rodents and shrews
Between 2012 and 2014, a total of 345 specimens were collected in Yue-xiu (n = 170) and Baiyun (n = 119) districts of Guangzhou city and Tong-an (n = 20) and Haicang (n = 36) districts of Xiamen city. Of the 209 samples collected from R. norvegicus, 16 (7.7%) tested positive for HVs, whereas no positive result was found for Rattus tanezumi (n = 9) and M. musculus Linnaeus (n = 10). One (3.7%) of the 27 samples from R. lossea Swinhoe tested positive for HVs. Six (6.7%) of the 90 S. murinus specimens yielded a positive result for HVs. All sites for collection tested positive for HVs (Fig. 1 and Table 1).
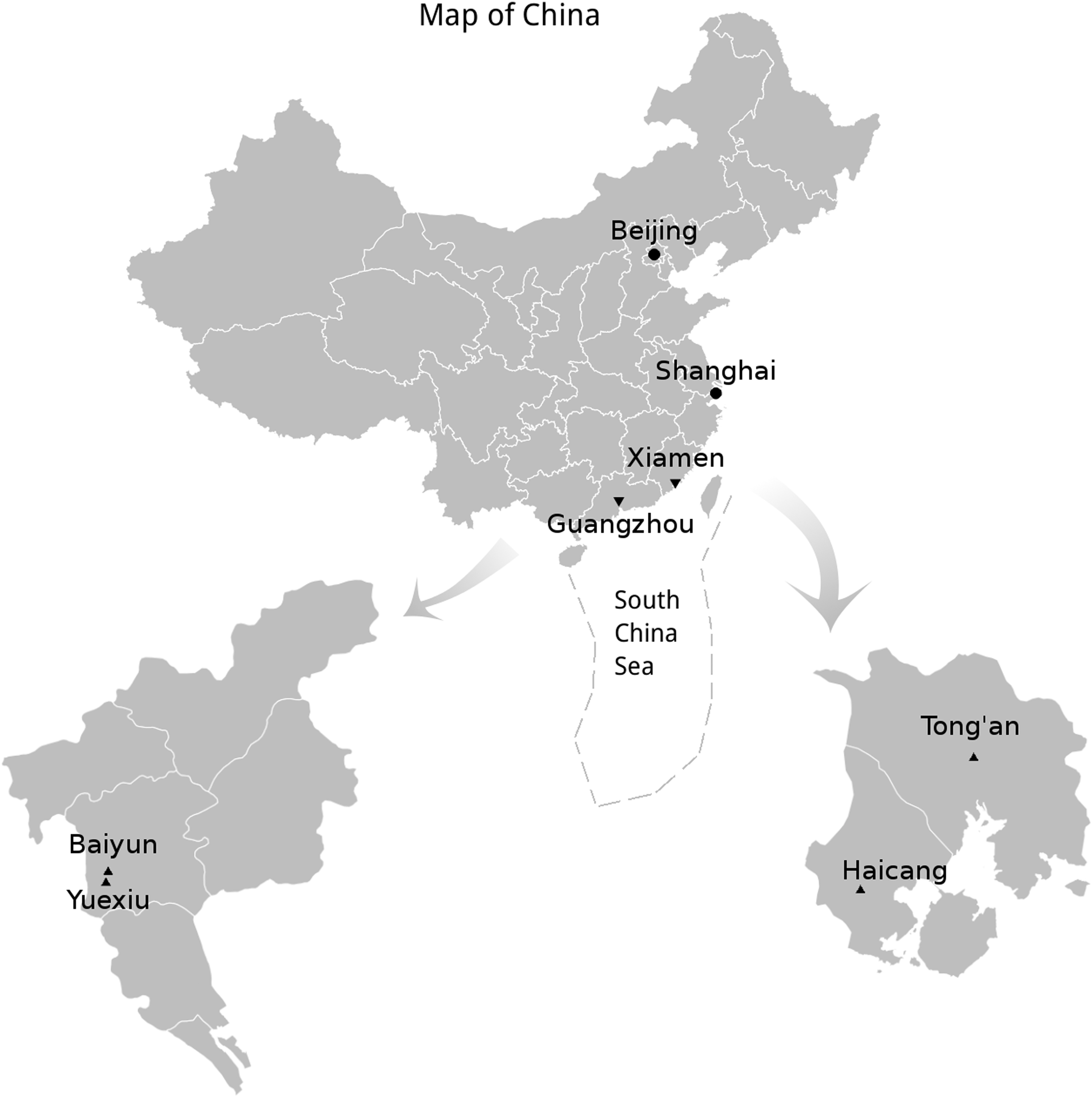
Geographical distribution of rodent samples in southern China. Triangles indicated the sampling locations.
GZYX, Yuexiu district in Guangzhou city (23.13°N; longitude: 113.27°W); GZBY, Baiyun district in Guangzhou city (23.17°N; longitude: 113.27°W); XMHC, Haicang district in Xiamen city (latitude: 24.47°N; longitude: 117.98°W); XMTA, Tong-an district in Xiamen city (latitude: 24.73°N; longitude: 118.15°W); No., the number of rodents and shrews collected; (+), the number of rodents/shrews detected positive for herpesviruses; positive %, the positive detection rate of herpesviruses in different rodent/shrew species.
Phylogenetic analysis of HVs
Phylogenetic analysis of the partial DPOL gene sequences revealed that rodent HVs were clustered into two subfamilies of Betaherpesvirinae (3 positive samples) and Gammaherpesvirinae (14 positive samples). All shrew HVs were clustered into Gammaherpesvirinae (six positive samples). None of the Alphaherpesvirinae subfamily was detected (Fig. 2).

Phylogenetic analysis based on 54 amino acids of the DNA-directed DNA polymerase (DPOL) of HVs detected from rodents in southern China. The tree was generated using the neighbor-joining method with the p-distance model. A bootstrap test was replicated for 1000 times. Numbers above the branches indicate NJ bootstrap values. Solid triangles indicate the positive HV samples of our present study. Solid squares indicate the bat HVs detected in the context of our study. RLS, Rattus lossea Swinhoe; RN, Rattus norvegicus; SM, Suncus murinus; YX, Yuexiu district; BY, Baiyun district; XM, Xiamen city; HV, herpesvirus.
Gammaherpesvirinae could be subdivided into six subgroups: G1 included the proposed Percavirus genus, detected in mustelids, bats, and equids; G2 contained the proposed Rhabdinovirus, detected in S. murinus, bats, humans, and bovine and ateline animals; G3 incorporated the proposed Lymphocryptovirus, detected in humans and macacine animals; while G4 included the proposed Macavirus, detected in rat, ovine, bovine, and alcelaphine animals. Distinct from the other genuses, the remaining two clades of Gammaherpesvirinae comprised prominent rodent-only clades, the phylogenetic placement of which in Gammaherpesvirinae was unclear, even in the phylogenetic tree with bootstrap value of 1000. In terms of the Betaherpesvirinae subfamily, three genuses could be observed in this clade: B1 (proposed as Roseolovirus, detected in humans), B2 (proposed as Cytomegalovirus, detected in rat), and B3 (proposed as Muromegalovirus, detected in rodent animals) (Fig. 2).
Sequence identities of the deduced amino acids of DPOL, identified among all rodents and shrews, which tested positive for HVs, revealed a significant divergence, which ranged from 42.3% to 100.0%. Interestingly, a higher diversity could be observed in rodent HVs (similarity 44.0–100.0%, with 81.0% of the similarity values being less than 90%) compared with S. murinus HVs (similarity 90.0–100%, with none of these similarity values being less than 90%). The similarities of R. norvegicus/R. lossea Swinhoe and R. norvegicus/S. murinus HVs were 44.0–71.9% and 45.7–70.1%, respectively (Supplementary Table S1; Supplementary Data are available online at
The herpesvirus DPOL sequences, found in 23 samples, could be classified into five clades. Of these, four clades were found in commensal rodents (R. norvegicus) and field rodents (R. lossea Swinhoe) in this study, which were identical to those known sequences of HVs from R. norvegicus (R. norvegicus rhadinovirus 1, Murid herpesvirus 8), Bandicota indica (Bandicota indica rhadinovirus 4), and bovine (Bovine lymphotropic herpesvirus) organisms (Supplementary Table S2, BLAST results). Six S. murinus (SM/14BY7/16/20/97/99/106) specimens tested positive for HVs and these HVs were apparently different from the existing rodent HVs, sharing only a 28.3–76.1% identity with the rodent DPOL sequences (values not shown). The deduced DPOL amino acid identity of HVs in the present study, compared with other original HVs, is shown in Supplementary Table S1. In the proposed phylogenetic tree of the DPOL gene, six S. murinus HVs had only 60–76.3% and 80.0% identity with BoHV-4 and AtHV-1, respectively. Notably, HVs from S. murinus (SM/14BY7/16/20/97/99/106) specimens shared identities of 42.5–75% with three bat HVs (SK/13YF239, RB/13YF104, MS/11HN96) in the context of our recently published findings of gammaherpesviruses in bats compared with the similarity of 44.4–100% (5% of the values were larger than 90%) between rats and bats. Interestingly, one bat HV (MS/11HN96) in our recent study shared 100% identity with BoHV-6 and four R. norvegicus HVs (RN/13YX23/24/52 and RN/14HC/50) compared with a 46.2–61.5% identity with other rat HVs in this study.
For amplification of gB gene sequences of the Betaherpesvirinae subfamily, only three specimens (RN/12YX60, RN/13YX28/31) tested positive. Phylogenetic analysis showed that these three specimens clustered with the previously reported murine HVs, constituting the Muromegalovirus, which shared a high similarity of the gB amino acids. Compared with other animals and human HVs, these three HVs—which belonged to Betaherpesvirinae—shared a 71.8–72.8% similarity to BBHV-2, 62.1–63.1% similarity to MaHV-3, 56.3–57.2% similarity to HHV-5, 57.5–58.4% similarity to HHV6, and 54.7–55.6% similarity to HHV7 (Fig. 3 and Table 2).
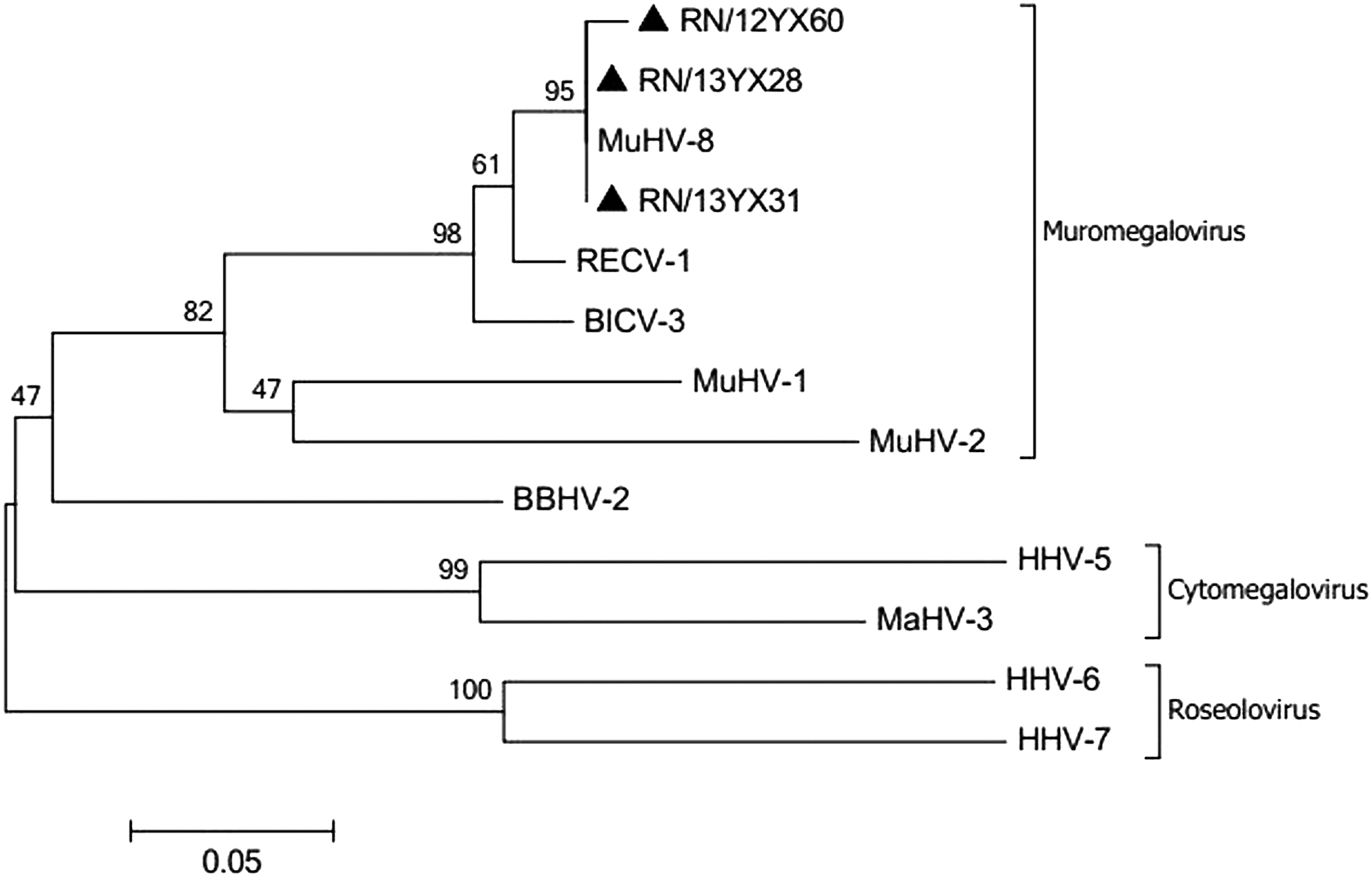
Phylogenetic analysis based on 106 amino acids of the (Betaherpesvirinae gB) of HVs detected from rodents in southern China. Solid triangles indicate the positive HV samples of our present study. Abbreviations of virus names are as shown in Figure 2, footnote.
BBHV-2: Bat betaherpesvirus 2, AB517983.1; BICV-3: Bandicota indica cytomegalovirus 3, EF125067.1; HHV-5: Human herpesvirus 5, M17209.1; HHV-6: Human herpesvirus 6, M97927.1; HHV-7: Human herpesvirus 7, D86577.1; MaHV-3: Macacine herpesvirus 3, NC_006150.1; MuHV-1: Murid herpesvirus 1, HE610456.1; MuHV-2: Murid herpesvirus 2, NC_002512.2; MuHV-8: Murid herpesvirus 8, KP202868.1; RECV-1: Rattus exulans cytomegalovirus 1, EF125071.1; RLS: Rattus lossea Swinhoe; RN: Rattus norvegicus; YX: Yuexiu district.
For a more meaningful phylogenetic analysis of Gammaherpesvirinae, a deg/dI nested primer set targeting the gB gene was used. All Gammaherpesvirinae HVs (20 specimens) detected by the partial DPOL gene were successfully amplified with the gB gene sequences. Most of the genus classification was consistent with the phylogenetic tree of the DPOL gene. Eight R. norvegicus HVs (RN/12YX3/8/12/29/23/50/89, RN/14YX4, and RLS/14XM17) were clustered with the previously reported murine HVs, while six shrew HVs (SM/14BY106/16/7/20/97/99) were clustered with bat HVs. Similar to the DPOL gene, the placements of the shrew HV branches could not be assigned, distinguishing them from the other genus group. In particular, six S. murinus HVs (SM/14BY7/16/20/97/99/106) shared identities of 46.9–61.7% with four bat gammaherpesviruses (HP/11HN110, HP/11HN104, RB/13YF87, RB/11HN56) compared with the similarity of 45.2–99.3% (5.4% of the values were larger than 90%) between rats and bats in southern China based on our recent findings of bat HVs. Two R. norvegicus HVs (RN/12YX23/8) were classified within the Gammaherpesvirinae subfamily based on the deduced gB amino acids, assigned as Betaherpesvirinae according to the DPOL gene classification. Three R. norvegicus HVs (RN/13YX52/24 and RN/14XM50) were clustered together with BLHV, BoHV-6, Rb/13YF87, AIHV-1, and OvHV-2, members of the Macavirus genus. Similarly, the bat HV (Rb/13YF87) in this clade shared identities of more than 95% with bovine herpesviruses 6 and three R. norvegicus HVs (RN/13YX24/52 and RN/14XM50) compared with 52.9–58.8% identities with other rodent HVs in this study (Fig. 4). Sequence identities of the Gammaherpesvirinae gB gene amino acids among rodent HVs reported in this study ranged from 53.6% to 100.0%, with 65.5% of the values being less than 90.0%. The percentages of identities to known HVs of Gammaherpesvirinae—including bat, bovine, human, ovine, alcelaphine, and murine animals—are shown in Table 3.

Phylogenetic analysis based on 152 amino acids of the (Gammaherpesvirinae gB) of HVs detected from rodents in southern China. Abbreviations of virus names are as shown in Figure 2, footnote. Solid triangles indicate the positive HV samples of our present study. Solid squares indicate the bat HVs detected in the context of our study.
AlHV-1: Alcelaphine herpesvirus 1, NC_002531; AFRV-1: Apodemus flavicollis rhadinovirus 1, DQ821580; AtHV-3: Ateline herpesvirus 3, NC_001987; BIRV: Bandicota indica rhadinovirus 4, EF128043.1; BoHV-4: Bovine herpesvirus 4, AF318573; BoHV-6: Bovine herpesvirus 6, NC_024303; EHV-2: Equid herpesvirus 2, HQ247792; ESRV-1: Eptesicus serotinus rhadinovirus 1, DQ788623; HHV-4, Human herpesvirus 4, LN827580; HHV-8, Human herpesvirus 8, GQ994935; MaHV-4: Macacine herpesvirus 4, NC_006146; MuHV-4, Murid herpesvirus, NC_001826; MustHV-1: Mustelid herpesvirus 1, AF376034.1; MyHV-1: Myotis ricketti herpesvirus 1, JN692429; OvHV-2: Ovine herpesvirus 2, NC_007646; RLS: Rattus lossea Swinhoe; RN: Rattus norvegicus; SM: Suncus murinus; YX: Yuexiu district; BY: Baiyun district; HC: Haicang district; TA: Tong-an district; XM: Xiamen city.
Discussion
Using PCR techniques that targeted two conserved genes, this study presented the results of an integrated search for alphaherpesviruses, betaherpesviruses, and gammaherpesviruses in R. norvegicus, R. tanezumi, R. lossea Swinhoe, M. musculus Linnaeus, and S. murinus in southern China. Based on 345 rectal swab samples with a positive detection rate of 6.7%, this study has identified the novel R. norvegicus and S. murinus HVs belonging to the Gammaherpesvirinae subfamily.
The studies conducted by Ettinger et al. (2012) and Teterina et al. (2009) have offered novel insights into rodent betaherpesviruses, which harbored a high similarity among the newly detected viruses in M. musculus and R. norvegicus. Likewise, the betaherpesviruses (RN/12YX60, RN/12YX28/31) found in this study were also highly similar to the published muromegaloviruses. In the first globally comprehensive search conducted by Ehlers et al. (2007), the positive detection rate was 27%, with the identification of 17 novel betaherpesviruses and 21 gammaherpesviruses. The comparatively low positive detection rate (6.7%) found in the rectal swab samples in our study most likely reflected the biology of the virus. The virus may reside in the spleens of all infected animals (Hsieh et al. 2010); therefore, only a small proportion of these viruses can be detected in the rectum.
The rat and shrew HVs detected in this study were found to be generally species specific based on the following evidence: (1) R. norvegicus and shrew HVs were phylogenetically distinct from the HVs derived from other animals; (2) low pairwise amino acid identities of different genes (DPOL and gB) could be observed between the HVs in this study and other original HVs, indicating that these rodent and shrew viruses were genetically distinct from the HVs of other species; and (3) a phylogenetic relationship was shown to exist among these rodent viruses, as supported by the high post-test probability in two conserved genes.
Notably, two novel gammaherpesviruses have been identified. One of these was found in S. murinus, which was detected for the first time. Compared with other reported rodent and nonrodent DPOL sequences, low amino acid identity could be observed. Only bat HVs (MyHV-1) are phylogenetically related, although distinct from S. murinus, sharing a sequence identity of 74.5–76.3% and 63.9–65.9% in the DPOL and gB gene, respectively. Despite these findings, only 42.5–75% and 41.9–61.7% DPOL and gB gene similarities could be found between S. murinus HVs reported in this study and bat HVs in the context of our recently published findings of gammaherpesviruses in southern China after excluding the probability of PCR and sample contamination. The other novel rodent gammaherpesviruses (RN/13YX52/24 and RN/14XM50) clustered with BLHV, BoHV-6, AIHV-1, and OvHV-2, consisting of members of the Macavirus genus. To our knowledge, HVs are generally species specific, with limited reports of interspecies transmission (Donovan et al. 2009). Interestingly, two bat HVs (MS/11HN96, RB/13YF87) from Macavirus fell within four R. norvegicus HVs (RN/13YX23/24/52 and RN/14XM/50) and bovine herpesviruses 6, sharing the similarities of 97.9–100%, suggesting the potential of interspecies transmissions. This might lead us to speculate that bat HVs (MS/11HN96, RB/13YF87) may have a broad spectrum of hosts. However, owing to limited sequencing data, we could not comment further on their evolutionary history. Further investigation into bovine HVs may help elucidate their origins.
We did not find positive results of HVs in R. tanezumi and M. musculus Linnaeus. This may have been due to limited sample sizes in this study. Hence, larger sample sizes with longer durations and more abundant sites of sample collection are warranted. Moreover, consistent with the findings of other studies (Ehlers et al. 2007, Teterina et al. 2009, Hughes et al. 2010, Ettinger et al. 2012), none of the alphaherpesviruses were found in the rodents. This may have been due to the possibly low prevalence (particularly in the study with limited sample sizes) and the low viral loads in the fecal or rectal swab samples. It is also possible that rodent alphaherpesviruses had not evolved or ceased to propagate earlier during the evolution of the HVs. Further studies are needed to clarify this issue.
In conclusion, we found that the rodent and shrew HVs detected in this study are species specific. Two new HVs, found in S. murinus and R. norvegicus, have expanded the taxonomy of rodent and shrew HVs. In addition, the HVs found in this study have contributed to our understanding of the phylogeny and biology of HVs in southern China.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (grant no. 81373051).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.