
Abstract
Backgrounds:
Lymphatic filariasis is principally caused by Wuchereria bancrofti, and Brugia malayi. The other two filarial nematode species, Brugia pahangi and Dirofilaria immitis, possibly cause human zoonotic diseases.
Methods:
We propose the development of a PCR assay linked with DNA pyrosequencing as a rapid tool to identify W. bancrofti, B. malayi, B. pahangi, and D. immitis in blood samples and mosquitoes. Primers targeting the fragment of the 5S ribosomal RNA and spliced leader sequences were newly designed and developed to identify these four filarial nematodes. Analytical sensitivity and specificity were evaluated.
Results:
Pyrosequencing determination of nucleotide variations within 36 nucleotides for B. malayi and B. pahangi, and 32 nucleotides for W. bancrofti and D. immitis is sufficient for differentiation of those filarial nematodes, and for detection of intraspecies genetic variation of B. malayi. This analysis could detect a single B. malayi, B. pahangi, W. bancrofti, and D. immitis microfilaria in blood samples.
Conclusions:
Overall, the PCR-linked pyrosequencing-based method was faster than direct sequencing and less expensive than real-time PCR or direct sequencing. This is the possibility of choice that can be applied in a high-throughput platform for identification and surveillance of reservoirs and vectors infected with lymphatic filaria in endemic areas.
Introduction
L
Lymphatic filariasis is categorized by the World Health Organization as one of the six diseases, which can be completely eliminated (Morbidity and Mortality Weekly Report 1993). In addition, the other species, Brugia pahangi and Dirofilaria immitis, infect animals in endemic areas of Southeast Asia (Orihel 1961, Denham and McGreevy 1977, Lim and Mak 1983) and possibly cause zoonosis (Palmieri et al. 1985, Simón et al. 2012).
To develop prevention and control strategies for the disease in human being and zoonosis in canine and feline reservoirs, improved diagnostic tests are needed for monitoring the presence of larvae in the vector and/or microfilariae in the final hosts. Microscopic analysis is the definitive detection method of parasite agents in mosquito vectors and/or in the blood samples. However, this method is laborious, requires highly experienced personnel, and exhibits a low level of sensitivity. A rapid immunochromatographic card test, which is more accurate to detect circulating antigen, is made available (Weil et al. 1997, McCarthy et al. 2012). Antibody detection assays have also been used, but results indicated past exposure rather than active infection (Rao et al. 2000, Vincent et al. 2000). In addition, antigen and antibody detection assays are hardly reliable for detection in mosquito vectors.
Molecular-based detection tools are regarded more precise since the methods ascertain recent infection and have been applied in several laboratory units for the reliable identification of filarial worms (Lizotte et al. 1994, Ramzy et al. 1997, Kluber et al. 2001, Chansiri and Phantana 2002, Fischer et al. 2002, Nuchprayoon et al. 2003, Fink et al. 2011, Latrofa et al. 2012). Seminested polymerase chain reaction-restriction fragment length polymorphism (Nuchprayoon et al. 2003) and PCR-based procedures have been used to amplify the DNA of B. malayi and B. timori (Lizotte et al. 1994, Kluber et al. 2001, Fischer et al. 2002, Latrofa et al. 2012) and Wuchereria bancrofti (Ramzy et al. 1997, Chansiri and Phantana 2002, Fink et al. 2011) from blood samples.
However, these methods require post-PCR processing such as agarose gel electrophoresis, which is time-consuming, and has restricted throughput, and a prejudice regarding carry-over contamination, as well as a tendency to produce spurious results. Currently, several molecular tests have been reported as diagnostic tools for the detection of these filarial worms, for example, loop-mediated isothermal amplification for detecting B. malayi and B. timori (Poole et al. 2012), real-time fluorescence resonance energy transfer PCR for detecting W. bancrofti (Lulitanond et al. 2004), B. malayi, B. pahangi (Thanchomnang et al. 2008, 2010a), D. immitis (Thanchomnang et al. 2010b), and Dirofilaria repens (Duscher et al. 2009), and high-resolution melting (HRM)-based real-time PCR for detecting B. malayi, B. pahangi, D. immitis, and W. bancrofti (Thanchomnang et al. 2013; Wongkamchai et al. 2015).
Pyrosequencing is a recently discovered DNA sequencing procedure that is done on the ascertaining of the released pyrophosphate during DNA synthesis (Ronaghi et al. 1996, 1998, Nyren 2007), and is well suited to large-scale screening of short-length DNA fragments. At this time, pyrosequencing has become an up-to-date parasitological approach for the species-level identification of Entamoeba (Stensvold et al. 2010). In addition, it has been used to differentiate several taxonomic groups of any rank, such as Plasmodium (Lulitanond et al. 2012), Paragonimus (Tantrawatpan et al. 2013), Trichinella species (Sadaow et al. 2015), human taeniid cestodes (Thanchomnang et al. 2014), and fishborne trematodes (Tantrawatpan et al. 2014).
Due to the fact that the pyrosequencing technique is a fast, technically simple, and high-throughput procedure, of which there has been no reported use for the identification of lymphatic filarial nematodes, this study aimed to establish a pyrosequencing technology for the differentiation and detection of B. malayi, B. pahangi, W. bancrofti, and D. immitis in blood samples and mosquitoes. The analytical sensitivity and specificity were also assessed.
Materials and Methods
Source of blood specimens and experimental infected mosquitoes
W. bancrofti-infected human blood samples (n = 1), B. malayi-infected cat blood samples (n = 1), and B. pahangi (n = 8) and D. immitis-infected (n = 5) dog blood samples, as well as W. bancrofti-infected Culex pipiens quinquefasciatus (n = 5), B. malayi-infected Aedes togoi (n = 5), and D. immitis-infected Aedes aegypti (n = 5) were received from frozen bank (−20°C) at the Department of Parasitology, Faculty of Medicine, Khon Kaen University. The details were previously described (Thanchomnang et al. 2013).
This study was approved by the Human Ethics Committee of Khon Kaen University, based on the Ethics of Human Experimentation of the National Research Council of Thailand (Reference No. HE42036), and by the Animal Ethics Committee of Khon Kaen University, based on the Ethics of Animal Experimentation of the National Research Council of Thailand (Reference No. 0514.1.12.2/50).). Informed consent was obtained from the patient.
Control samples for testing analytical sensitivity and specificity
For the specificity evaluation, healthy blood specimens from dogs (n = 5), cat (n = 1), and humans (n = 1) were obtained as negative controls. In addition, the other hemoparasite-infected samples consisting of human blood infected with Plasmodium falciparum (n = 1) and Plasmodium vivax (n = 1), as well as of dog blood infected with Hepatozoon canis (n = 1) and Babesia spp. (n = 2), were used for specificity testing. For sensitivity evaluations, each blood sample that was given a serial number of 1, 2, 4, and 8 of B. malayi and W. bancrofti microfilariae in human blood, of B. pahangi microfilariae in cat blood, and of D. immitis microfilariae in dog blood was used.
DNA preparation and primer design
Each mosquito was homogenized before DNA extraction using a NucleoSpin Tissue Kit (Macherey-Nagel). For blood samples, DNA was extracted from 50 μL of blood specimens using a NucleoSpin Blood kit (Macherey-Nagel). The adsorbed DNA was removed by washing with 100 μL of 5 mM Tris-HCl (pH 8.5) and was kept at −20°C until use.
The available nucleotide sequences surrounding the trans-spliced leader SL1 exon and 5S ribosomal RNA (rRNA) genes of B. pahangi (GenBank acc. no. D87038), B. malayi (GenBank acc. no. D87037), W. bancrofti (GenBank acc. no. AF297119), and D. immitis (GenBank acc. no. D87039) were selected and used to design the primer set based on the conserved region. This primer set (Proligo) was composed of forward primer (SLX-F: 5′-GGA ATC CCA GGT GTT GTA GAC AT-3′) and biotinylated reverse primer (SLX-R_Rb: 5′- Biotin-GGG CTG AAA CAT TCA ATT ACC TC-3′) for PCR, and sequencing primer (SLX-S: 5′-CCC AGG TGT TGT AGA CAT-3′) for pyrosequencing. All quantitative parameters of the primer set were evaluated for pyrosequencing with PyroMark Q96 ID software version 2.0 (Biotage). The multiple alignment of this primer set is presented in Figure 1.
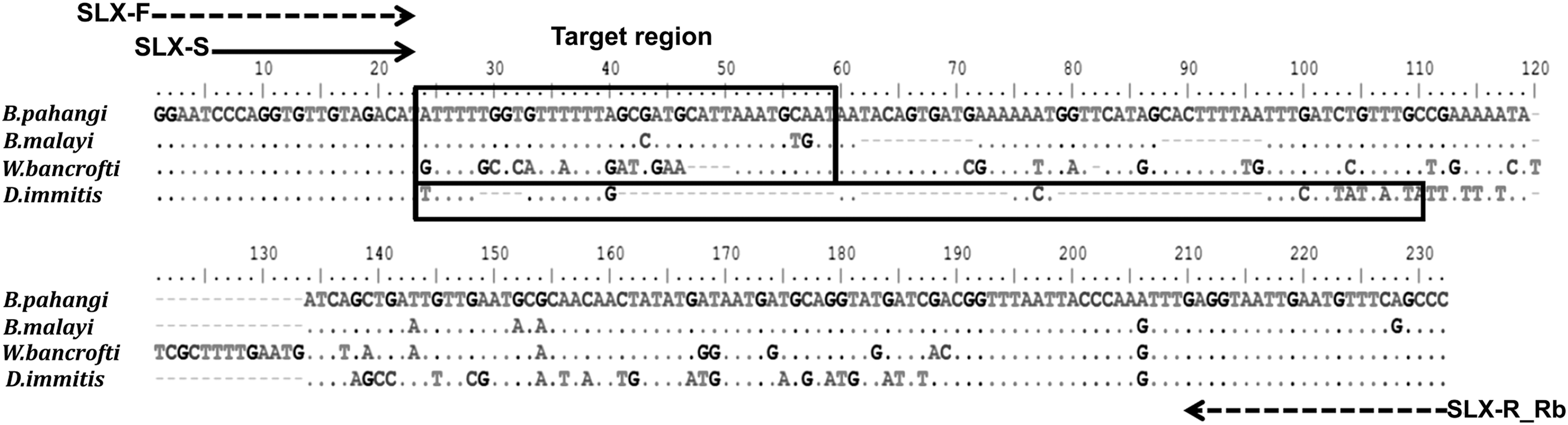
Multiple alignment of the sequences locating the trans-spliced leader SL1 exon in the 5S ribosomal RNA (rRNA) gene spacer region of Brugia pahangi, Brugia malayi, Wuchereria bancrofti, and Dirofilaria immitis. The dotted arrows show the positions of the forward primer (SLX-F) and biotinylated reverse primer (SLX-R_Rb). The solid arrow shows the positions of the sequencing primer (SLX-S). The rectangles indicate the target regions for identification of filarial worms. Dots represent identical nucleotides between lines. Dashes indicate gaps between nucleotides.
PCR and pyrosequencing
According to a bioinformatics examination, the primer set can hybridize with W. bancrofti, B. malayi, B. pahangi, and D. immitis genes, yielding amplified products of different sizes. Positive control plasmids of individual species were constructed by amplicons using SLX-F and nonbiotinylated reverse (SLX-R) primers. They were ligated and transformed into pGEM®-T Easy vectors (Promega) and an Escherichia coli JM109, respectively. For validation of sequence accuracy, the nucleotides in inserted genes were bidirectionally sequenced by the Sanger method.
The reactions to PCR amplification were operated in a total volume of 25 μL containing 1× PCR buffer (Invitrogen) with 0.2 mM of each dNTP, 2 mM MgSO4, 0.4 μM of each primer, 0.625 U of Platinum Taq DNA polymerase high fidelity (Invitrogen), and 2 μL of the DNA specimen. The PCR assay was done using a GeneAmp PCR system 9700 thermal cycler (Applied Biosystems). The amplification process was as follows: 5 min at 94°C for initial denaturation followed by 35 cycles of denaturation for 30 s at 94°C, annealing for 30 s at 59°C, and extension for 30 s at 72°C, followed by a final extension for 7 min at 72°C. PCR amplicons were revealed using 1.5% agarose gel electrophoresis. The specific amplicon products that filled biotinylated primers were then done before pyrosequencing.
The pyrosequencing methods were performed by using the PyroMark™ Q96 ID instrument (Biotage) according to the manufacturer's protocols. The pyrosequencing procedure was followed that previously described (Tantrawatpan et al. 2013) with the exception that 0.4 μM-specific sequencing primer (SLX-S) was performed before doing the reaction. To evaluate the accuracy of the pyrosequencing results, all amplicons of each sample with SLX-F and nonbiotinylated SLX-R primers similarly with the above process were sequenced by the Sanger technique and compared with the pyrosequencing data.
Results
PCR amplification of sequencing templates
DNA from B. malayi-, W. bancrofti-, D. immitis-infected mosquitoes, and B. pahangi, W. bancrofti, D. immitis in blood samples, as well as from the positive control plasmids of each species, was amplified. The expected sizes were 199 bp for B. malayi, 218 bp for B. pahangi, 227 bp for W. bancrofti, and 164 bp for D. immitis. DNA from uninfected dog, cat, human blood samples as negative controls, and blood samples infected with other hemoparasites were not amplified by this SLX-F and SLX-R_Rb primer set (data not shown).
Pyrosequencing for the differentiation of filarial nematodes and its evaluation
The nucleotide variations within 36 nucleotides (for B. malayi and B. pahangi) and 32 nucleotides (for W. bancrofti and D. immitis) of 5S rRNA gene and spliced leader sequence SL1 close to the 3′ end of sequencing primer (SLX-S) were useful as the target region for the species differentiation of these filarial nematodes. The nucleotide sequences at the objective region of each filarial nematode were revealed as the pyrogram results (Fig. 2). At the target region, B. malayi could be discriminated from B. pahangi by three nucleotide positions (C43G, T56C, and G57A; Fig. 1), whereas W. bancrofti and D. immitis have several unique nucleotide sequences for each species identification (Figs. 1 and 2).

Representative pyrosequencing data of B. pahangi, B. malayi, W. bancrofti, and D. immitis included the pattern of pyrogram (top panel) and a raw data (bottom panel). Pyrosequencing based on the 5S rRNA gene and spliced leader sequence SL1 was done by the adding of four different nucleotides, enzyme (E) and substrate (S). The alphabets below the black bars represent the dispensation (Disp:) order. The pyrosequencing sequence is shown below the X-axis panel after “Seq:.” The level of fluorescence (Y-axis) emitted by the adding of a nucleotide base and the total number of bases (X-axis) put at that point; C, A, G, T, nucleotide bases were shown. Light gray areas indicate nucleotide positions used for differentiation between B. malayi and B. pahangi.
All samples and positive control plasmids of B. malayi, B. pahangi, W. bancrofti, and D. immitis exhibited the same sequence in the GenBank database as above except for one cytosine-to-adenine (C→A) substitution of B. malayi that was found (Fig. 3). The result accuracy was re-evaluated by Sanger sequencing of the amplicons, which revealed the same sequence data. For comparative efficacy, same samples were also performed by microscopy and Sanger sequencing. The pyrosequencing results were similar to the sequences showed by Sanger sequencing, which coincided with our morphological microscopic examination (Table 1).
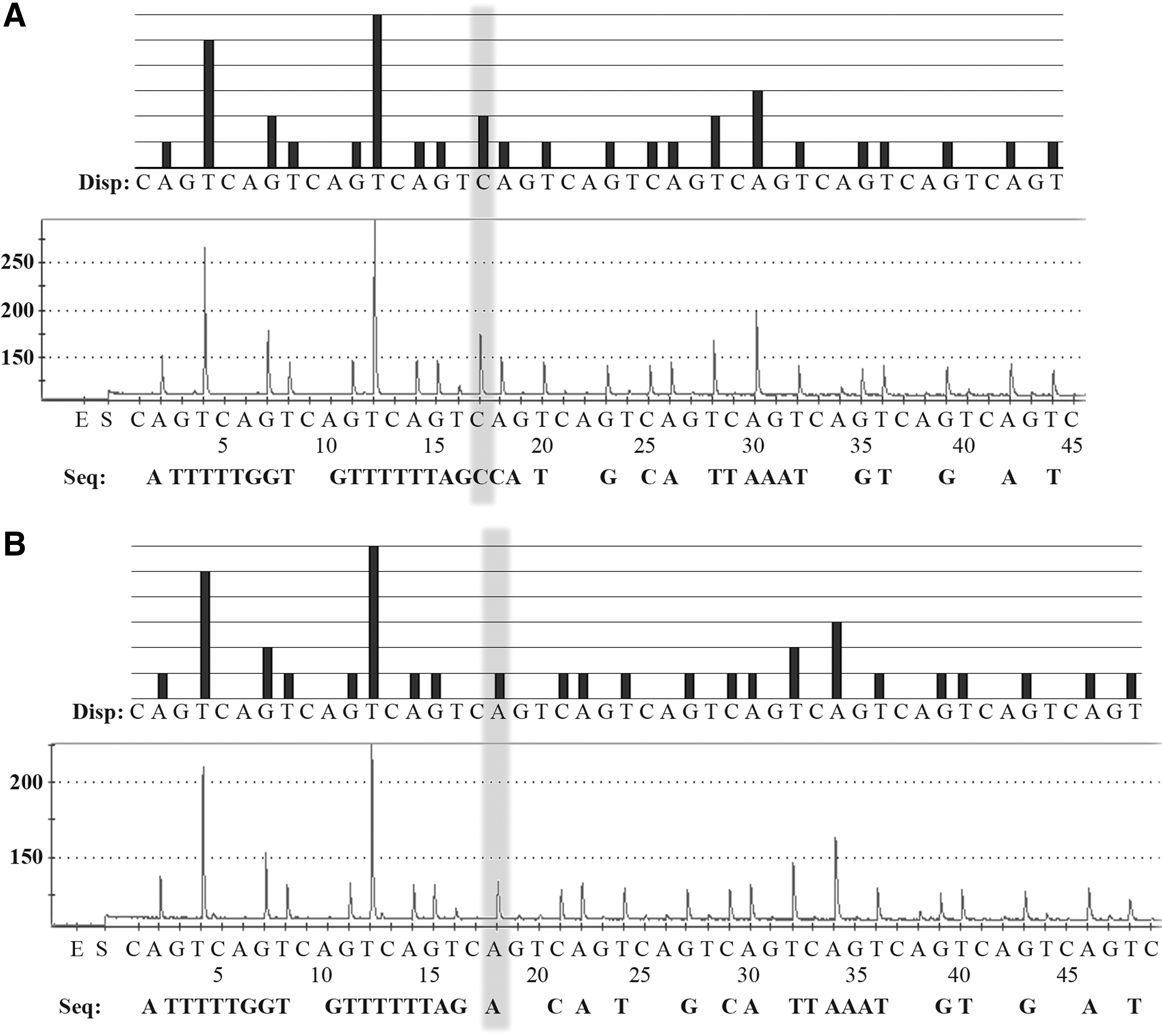
Pattern of pyrogram (top panel) and a raw data (bottom panel) of B. malayi. A single-nucleotide variation, exhibited as light gray, was evaluated to distinct cytosine
Standardization of pyrosequencing assay
To demonstrate the specificity of the pyrosequencing approach, two parallel control experiments that included only the sequencing primer or only the biotinylated primer was performed. Pyrosequencing results of other hemoparasites, that is, P. falciparum, P. vivax, H. canis, or Babesia spp. and all negative controls showed no pyrogram (data not shown). The analytical sensitivity enabled the identification of as little as a single microfilaria of D. immitis, B. malayi, B. pahangi, and W. bancrofti in a 50-μL blood sample.
Discussion
In the operating system of pyrosequencing, two modes, sequence analysis (SQA) and single-nucleotide polymorphisms (SNP), are available for use and each is able to detect different aspects. SQA mode ascertained short- and medium-length DNA fragments, and SNP mode ascertained only a single base variation. We developed a tool for the differentiation of D. immitis, B. malayi, B. pahangi, and W. bancrofti using the pyrosequencing technology with the SQA mode and assessed this analysis on a panel of blood samples and infected mosquitoes in comparison with Sanger sequencing and microscopic examination.
The specific primer set (SLX-F and SLX-R_Rb for PCR and SLX-S for pyrosequencing) was constructed depending on the conserved nucleotide areas in a consensus sequence from the 5S rRNA gene and spliced leader sequence SL1, which are conserved in B. malayi, B. pahangi, and D. immitis, although other areas of the spacer region are variable in length and sequence (Harasawa et al. 1997).
Bancrotian and brugian filariasis are important mosquito-borne diseases in tropical countries, including Thailand (World Health Organization 1992, Triteeraprapab et al. 2001). A prevalence as high as 4.4% was reported in Myanmar immigrant workers in Tak province, in northern Thailand (Triteeraprapab and Songtrus 1999), which could possibly lead to an outbreak of this harmful disease in Thailand. Highly specific and sensitive evaluated methods for monitoring are essential for the elimination program of the disease. This prompted us to develop the pyrosequencing assay for detection and differentiation of four filarial species; B. pahangi, B. malayi, W. bancrofti, and D. immitis in blood samples and mosquitoes.
In this study, the short (36 nucleotides for B. malayi and B. pahangi and 32 nucleotides for W. bancrofti and D. immitis) target regions in the 5S rRNA gene and spliced leader sequence SL1 were useful for species differentiation using pyrosequencing. This result is supported by the previous report that the sequence is a multicopy within the worm genome and can be used for the specific and sensitive detection of W. bancrofti (Dassanayake et al. 2001). By using this target region as the marker, B. malayi is distinct from B. pahangi by three nucleotides at the position nos. 43 (C→G), 56 (T→C), and 57 (G→A) (Fig. 1). Moreover, W. bancrofti and D. immitis sequences differ from others at many positions (Figs. 1 and 2). This result can be easily used for diagnosis among these filarial worms.
When PCR-amplified products were sequenced by the Sanger technique and evaluated with the pyrosequencing results, both sets were similar to each other at the destination region. The results showed the good reliability of pyrosequencing data. Furthermore, the primer pair did in our study reveal high specificity, as no amplicons were found with control genomic DNA, including DNA from the other hemoparasites. Handing pyrosequencing, many samples can be done in a short time compared with Sanger sequencing, at much cheaper expense for reagents with the same degree of accuracy (Fakruddin et al. 2012).
The usefulness of our pyrosequencing assay is its sensitivity. This assay can detect either a single B. malayi or W. bancrofti microfilaria in human blood as it can measure about 100 pg of DNA per microfilaria (Lizotte et al. 1994, Rao et al. 2006). This result is similar with the limit of detection of either B. malayi or W. bancrofti single microfilaria in blood using HRM real-time PCR (Thanchomnang et al. 2013). Therefore, the test could possibly be useful for diagnosis of B. malayi or W. bancrofti in human blood samples and monitoring of their transmission in mosquito vectors. Moreover, for future differentiation of mixed filarial cases, SNP pyrosequencing or uniplex SNP pyrosequencing and PCR amplification can be used to ascertain each base variant in the target region (Marsh 2007, Dabrowski et al. 2012).
In addition, the B. malayi (originating from Narathiwat, Thailand) sequence in mosquitoes from this study showed the C → A substitution (Fig. 3), which differs from the B. malayi originating from the Maeda strain in the GenBank database (GenBank acc. no. D87037), indicating the method can be applied for studying B. malayi genotyping or genetic diversity among a population. The use of the drugs could alter the genetic materials (e.g., SNP or mutations) of the parasites, leading to drug resistance as evidenced by reports regarding tuberculosis. Pyrosequencing could potentially be applied as a diagnostic tool for screening or predicting resistant strains (Zheng et al. 2014).
Conclusions
This study showed that pyrosequencing is a fast, accurate, cost-effective, and high-throughput assay for the differentiation and detection of four species of lymphatic filarial worms in both blood samples and mosquito vectors. However, larger sample numbers and more different isolated samples or varied parasite strains in endemic areas are needed to evaluate this assay as the further step in development regarding it being applied as a field tool for diagnosis of human blood specimens, molecular xenomonitoring in vectors, or detecting elimination in transmission after giving mass drug processing.
Footnotes
Acknowledgments
This work was funded by grants from the Higher Education Research Promotion of National Research University Project of Thailand, Office of the Higher Education Commission through the Health Cluster (SHeP-GMS); the Faculty of Medicine (Grant no. TR57201) and Post Doctoral Training Program of Graduate School and Khon Kaen University (Grant no. 58101), Khon Kaen University; and the Thailand Research Fund Senior Research Scholar Grant (Grant no. RTA5880001). The authors acknowledge the support of the English consultant Clinic, Research Affair, the Faculty of Medicine, Khon Kaen University and the Khon Kaen University Publication Clinic, Research and Technology Transfer Affairs, Khon Kaen University, for their assistance.
Author Disclosure Statement
No competing financial interests exist.